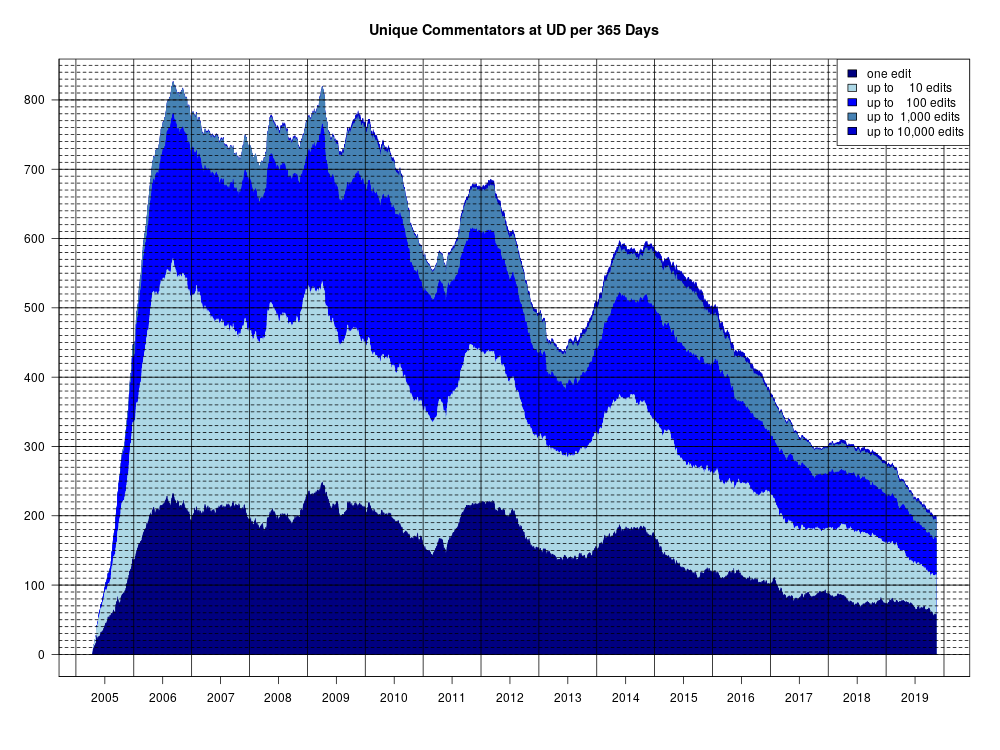
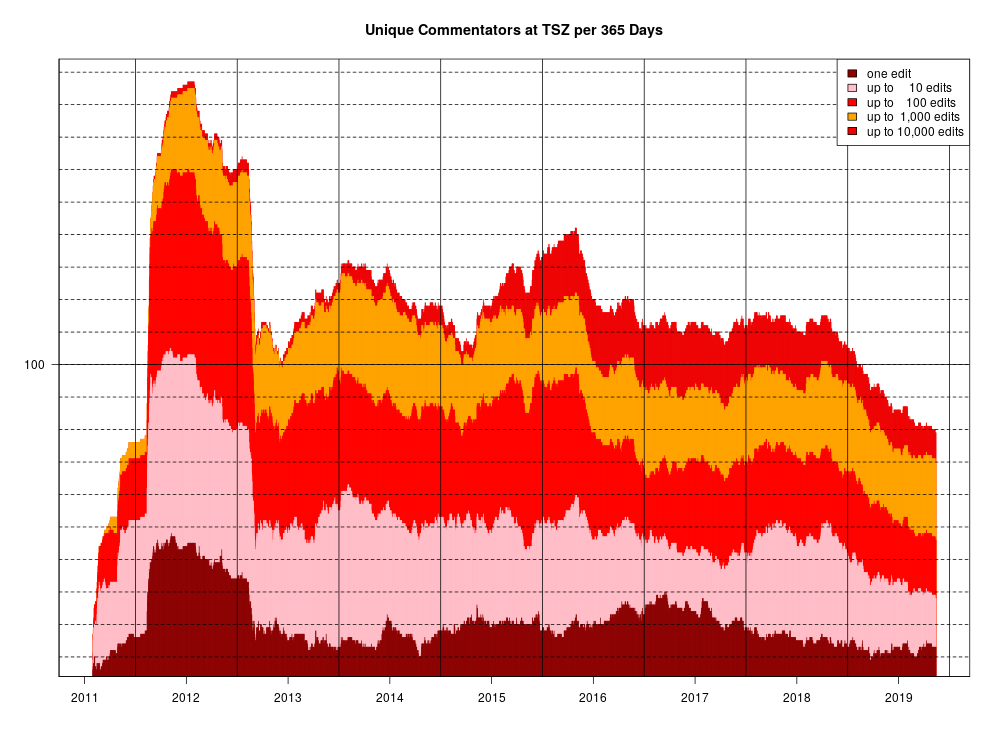
At Uncommon Descent, poster gpuccio has expressed interest in what I think of his example of a safecracker trying to open a safe with a 150-digit combination, or open 150 safes, each with its own 1-digit combination. It’s actually a cute teaching example, which helps explain why natural selection cannot find a region of “function” in a sequence space in such a case. The implication is that there is some point of contention that I failed to address, in my post which led to the nearly 2,000-comment-long thread on his argument here at TSZ. He asks:
By the way, has Joe Felsestein answered my argument about the thief? Has he shown how complex functional information can increase gradually in a genome?
Gpuccio has repeated his call for me to comment on his ‘thief’ scenario a number of times, including here, and UD reader “jawa” taken up the torch (here and here), asking whether I have yet answered the thief argument), at first dramatically asking
Does anybody else wonder why these professors ran away when the discussions got deep into real evidence territory?
Any thoughts?
and then supplying the “thoughts” definitively (here)
we all know why those distinguished professors ran away from the heat of a serious discussion with gpuccio, it’s obvious: lack of solid arguments.
I’ll re-explain gpuccio’s example below the fold, and then point out that I never contested gpuccio’s safe example, but I certainly do contest gpuccio’s method of showing that “Complex Functional Information” cannot be achieved by natural selection. gpuccio manages to do that by defining “complex functional information” differently from Szostak and Hazen’s definition of functional information, in a way that makes his rule true. But gpuccio never manages to show that when Functional Information is defined as Hazen and Szostak defined it, that 500 bits of it cannot be accumulated by natural selection.
Continue reading →