
Evolution of KRAB Zinc Finger Proteins vs. the Law of Large Numbers
There are patterns in biology that violate the law of large numbers, and thus suggests Intelligent Design or at the very least statistical miracles. The pattern involves KRAB-ZnF proteins that have multiple zinc finger domains side by side that are inexact copies of each other and would require a scenario of co-evolution of their DNA binding partners with every additional zinc-finger insertion — a scenario indistinguishable from a miracle.
The role of a zinc finger can be something like a clamp or a lock. Each zinc finger in a KRAB-ZnF protein is fine-tuned to connect with a DNA much like a lock (the zinc finger) can receive a key (like DNA). Here is a depiction of a KRAB-ZnF protein with 4 zinc fingers as part of a large chromatin modifying complex. The four zinc fingers are marked “ZN” and connect to DNA:
The evolutionary explanation of side-by-side repeated patterns of zinc fingers such as in KRAB-ZnF proteins shows a severe lack of critical thinking by evolutionary biologists who pretend “phylogenetic methods” are adequate explanations of mechanical feasibility of common descent.
To illustrate the problem, consider the KRAB-ZnF protein known as ZNF136. For reference, this is the amino acid sequence of ZNF136:
https://www.uniprot.org/uniprot/P52737.fasta
This is an amino acid fragment contained within the ZNF136 protein
TGEKLYDCKECGKTFFSLKRIRRHIITH
This short sequence is called a zinc finger domain which in 3D looks like this:
https://sciencescienceeverywhere.files.wordpress.com/2015/08/figure-11.jpg
{kind=link}
Wiki gives a description of the function of zinc fingers in proteins that have them:
http://wikipedia.org/wiki/Zinc_finger
A zinc finger domain requires two “C” amino acids and two “H” amino acids placed in the right positions. It requires a few other things too…
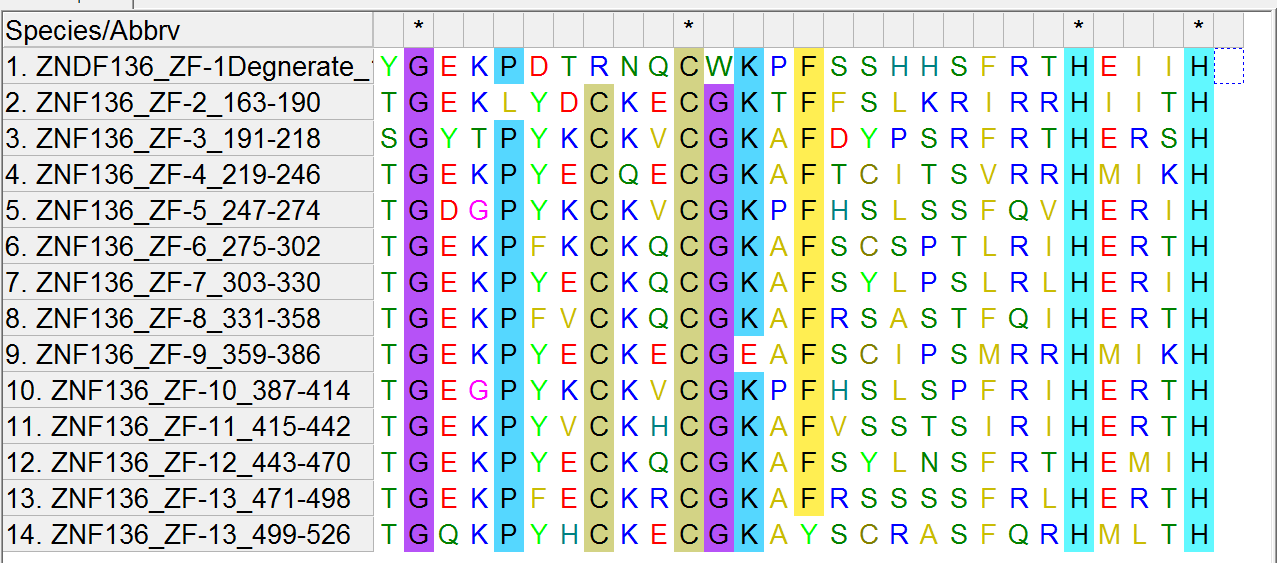
There are 13 zinc fingers in the ZNF136 protein and these are their sequences:
TGEKLYDCKECGKTFFSLKRIRRHIITH
SGYTPYKCKVCGKAFDYPSRFRTHERSH
TGEKPYECQECGKAFTCITSVRRHMIKH
TGDGPYKCKVCGKPFHSLSSFQVHERIH
TGEKPFKCKQCGKAFSCSPTLRIHERTH
TGEKPYECKQCGKAFSYLPSLRLHERIH
TGEKPFVCKQCGKAFRSASTFQIHERTH
TGEKPYECKECGEAFSCIPSMRRHMIKH
TGEGPYKCKVCGKPFHSLSPFRIHERTH
TGEKPYVCKHCGKAFVSSTSIRIHERTH
TGEKPYECKQCGKAFSYLNSFRTHEMIH
TGEKPFECKRCGKAFRSSSSFRLHERTH
TGQKPYHCKECGKAYSCRASFQRHMLTH
For completeness, there is a degenerate zinc finger in ZNF136 with the sequence “YGEKPDTRNQCWKPFSSHHSFRTHEIIH”
Why are the Zinc Fingers so different in sequence (except for the conserved amino acids)? To target a section of DNA, the zinc finger must be tuned to target it. Think of the zinc finger like a lock and DNA as a key that fits into the lock. In fact, for both the study of biology and medical applications, humans have a desire to make their own zinc fingers — like lock smiths. To bind a large segments of DNA, side-by-side zinc-fingers have to be tuned to their respective side-by-side DNA partners such as illustrated here:
By the way, there is a website that helps researchers construct the right amino acid sequence to make a zinc finger for a particular DNA target:
https://www.scripps.edu/barbas/zfdesign/zfdesignhome.php
Now, to visualize the critical/conserved amino acids, see the protein sequence here with highlights on “C” and “H” amino acids.
http://theskepticalzone.com/wp/wp-content/uploads/2019/04/znf136_zfC2H21.png
{kind=link}
Note one of the lines is not exactly like the other lines in that it is missing a “C”. This is the degenerate zinc finger mentioned above. So there is 1 degenerate zinc finger and 13 functional ones.
From this diagram it is apparent that the regular appearance of “C” and “H” is a violation of the law of large numbers, hence this pattern is not due to random point mutation alone. To “solve” this problem, Darwinist explain the pattern through segment duplication followed by some point mutation and natural selection, but this is not mechanically feasible either!
To understand why, let the reader first ponder the alignment I made of the Zinc Fingers in the ZNF136 protein using MEGA 6.0/MUSCLE software:
http://theskepticalzone.com/wp/wp-content/uploads/2019/04/znf136_zfC2H2_muscle1.png
{kind=link}
Then let the reader, consider the distance matrix generated by MEGA 6.0 which measures the number of nucleotide and percent differences between the zinc fingers.
http://www.creationevolutionuniversity.org/public_blogs/reddit/znf136_distance_matrix.xls
(All of the above results are reproducible, so I leave it to interested parties wanting to confirm the results to do so.)
For the duplication to work, at a bare minimum the right 84 nucleotide segment must be chosen, and then perfectly positioned for insertion so as not to break a pre-existing zinc finger. But supposing the duplication succeeds, why are the zinc finger’s conserved features involving “C” and “H” and other amino acids preserved and not eventually erased by point mutation given we obviously see the zinc fingers are different from each other. To preserve the “C” and “H” and other necessary amino acids in a zinc finger, the new zinc finger needs to be under selection. But in that case one is simply concocting a “just so” story for those newly minted zinc fingers without any respect for the difficulty of such a “just so” story being probable — and it is not probable!
To understand the problem of such a “just so” story, recall zinc fingers bind to DNA regions. Btw, this includes DNA regions such as ERVs! And ERVs are indicated to participate in the Stem Cell Pluripotency regulatory network:
https://www.nature.com/articles/nature13804
Further, the KRAB-ZnF protein is part of an incredibly complex machine that does chromatin modification (as shown above) by often attaching to ERV targets. But this would require that ERVs (or whatever DNA target) needs to co-evolve with the KRAB-ZnFs that attach to them!
Do these researchers even consider the fact such evolution would have to be instantaneous otherwise it would degrade function and not let the genome have the opportunity to adapt to the new accidental copy of the zinc finger because the duplication would immediately be selected against!
The above KRAB-ZnF complex is like a read/write head acting on Chromatin. Chromatin itself is an amazing mind-boggling design akin to computer ROM and RAM in one.
Again, the difficulty of evolution via random insertion/duplication mutations followed by point mutations is that such events would disrupt the binding of an already operational set of zinc fingers. For example, suppose we have an array of 10 zinc fingers side-by-side that collectively bind to a target DNA. Suppose one zinc-finger is duplicated and the number of zinc fingers is increased from 10 to 11. Oh well, the binding ability is broken or at best compromised, much like adding a single letter to a pre-existing password!
In sum, there is a violation of the law of large numbers in KRAB-ZnF proteins which is not explained by random mutation, nor random segment duplication followed by some point mutation and fixed by natural selection. Some other mechanism for the emergence of such proteins is indicated and would likely be indistinguishable from a miracle. Given the importance of such zinc finger proteins in the control of ERVs which are important in the stem cell pluripotency regulatory circuits, the origin of KRAB-ZnFs is even more miraculous.
See my OP on the binding affinity aspect of your failure to comprehend.
The “random ectopic” thing is dumb beyond belief; again, if anyone-but-Sal has any doubts on that subject, let me know…
Sal rants
No Sal, that’s what I meant when I wrote
Sal attacks his own position
Good grief, no! That’s what YOU are saying. I disagree.
YES! That’s right!
“SHAZAM” is a great argument.Your analogy breaks down, however, because they spend most of their time NOT in a house. There is an “off” rate (unless you are streptavidin, which you are not).
Bill,
Now back to your questions. To introduce the zinc finger, let me point to something you might be familiar with, the Vitamin D Receptor:
https://www.uniprot.org/uniprot/P11473
It happens to have 2 zinc fingers. Below is a screen capture with the zinc finger data highlighted in red. I will describe more in subsequent comments.
Bill,
DNA not only serves to provide blueprints to proteins, but it can act as a scaffold/parking lot for molecular machines. The most amazing thing from ENCODE 2015 was fact the Vitamin D Receptor could park on introns, exons of on gene in order to regulate other genes!
So if we look at DNA as a parking lot, because each location has sequences associated with it, the sequences can act as a bar code of sorts to identify the parking lots in DNA. The zinc fingers are like bar code readers. When the right bar code is found, the molecular machine can park there (a machine like a Vitamin D receptor or Transcription Factor or whatever).
An individual zinc finger can target 3 or 4 bases. To get a really specific bar code, one needs multiple zinc fingers. I provided an example earlier for mouse ZFP932 that uses 4 zinc fingers to get this “bar code”:
Bill,
By way of contrast this is the Vitamin D Receptor binding logo that is presumably the target of the 2 zinc fingers on the Vitamin D Receptor-RXR complex:
Readers are invited to see the paper I linked to and decide for themselves. The time bound to unrelated sequences is called SCANNING in the paper, as in bumping into other sequences until Mr. Right House DNA is found.
In any case, I thank Jock for keeping my thread alive. I plan to post more in this thread, and he’s going to volunteer, I can see, to keep this from being a monologue — aren’t you. What a pal.
I used the word RELATIVE affinity several times, and DNA Jock chose to totally ignore it and misrepresent what I said. Here is an example of that:
In any case, relative affinity can be used as a measure of specificity. That was done so in this 1989 paper using affinity chromatography:
https://www.ncbi.nlm.nih.gov/pubmed/2674675
But now DNA jock has gone on record as saying and Identical tandem zf-repeat in a hypothetical ZFP protein would not change relative affinity of the by the new ZFP toward regions the ZFP bound to before the tandem repeat event.
Anyway, I know now how I’ll be mis-represented if I use the word (relative) affinity rather than specificity.
So thanks to DNA_jock for the conversation and showing how he intends to distort what I say. That was valuable input. I’ll have a better choice of words in my presentation.
Who will you be presenting your thoughts on zinc fingers to?
Suppose I’m God. I want to generate a TF composed of multiple units because … well, I’m God, do I have to explain everything? Having done so, I discover that I have unavoidably changed the binding characteristics relating to other sequence, because for some reason I’ve decided to constrain everything by the same basic forces. The more subunits I add, the greater the issue, which is a bit of a bugger because I want more subunits, dammit!
What have I done to get round this?
Consider the problem of a SOLITARY zinc-finger trying to bind to DNA. The solitary zinc finger can target 3 or 4 bases.
This means can pick out at best a proportion of 1 out 256 = 0.4% possible locations on a some arbitrary genome? So in a genome of 3.3 gigabases, it might pick out about 13 million sites? Not good. Not much for selection to work with.
Ok, so a TANDEM array, which will identify 6 to 7 bases. Better. 1 out of 4^6 = 1 out of 16384 = 1 out of 4096
So it might pick out 805,000 sites on the human genome. Still not too discirminating. So lets try 12 base identification. Good now we’re down to about 200 locations on the genome.
The point? Not much reason to believe even as single random tandem pair of zinc fingers will be functional for a transcription factor, much less a SOLITARY zinc finger. So then where did the first KRAB zinc finger come from. PLEASE no phylogeny BS, how about a description of evolution in terms of functionally feasible steps, not a phylogenetic hand wave.
A private group. I met some of the faculty in this presentation which was public which I announced at TSZ earlier:
Regarding KRAB ZFP and alternative splicing!!! YAY!
Yeah, now we’re starting to see what alternative splicing actually does for the organism!
https://www.ncbi.nlm.nih.gov/pmc/articles/PMC3731817/
DNA_Jock ignored the fact I used the word RELATIVE when describing affinity.
He wants to bash something other than the intended usage of my words, that’s on him, it doesn’t refute my point. He wants to change the meaning of what I’m claiming by carefully omitting my use of the word RELATIVE, that’s on him.
Here is a paper that uses the concept of relative affinity (expressed in terms of KDns/KD ratios) to describe specificity
https://www.pnas.org/content/95/6/2812
That’s exactly what I was getting at with my #25 example above becoming more specific than the other 63 targets after the “random” tandem repeat changes the zinc finger protein.
Additionally there is evidence the concatenated DNA string, as I had shown, is a good approximation of what the tandem zinc finger will bind to vs. the solitary zinc finger provided the solitary one is of good quality in specificity. This paper says as much:
https://genome.cshlp.org/content/26/12/1742.full
So DNA_Jock, refuted again. 🙂
That’s a good question. Of course, KRAB-zinc finger proteins have not evolved in the context of the human genome, but are suggested to have evolved as repressors of parasitic transposable elements in the vertebrate lineage.
I haven’t finished reading the paper, but this part seems important to your question at least:
I don’t know how plausible this scenario is, but at least it demonstrates that there are situations in which changes in (relative) binding affinity are not necessarily “bad juju”. You simply don’t know whether mutations will turn out to be deleterious or not.
Ouch, broken link. Here is the paper.
I am fairly sure that I have not gone on record saying anything that incoherent.
🙂
Please slow down and read for comprehension.
I am “carefully omitting” nothing, Sal.
Your recollection of the conversation is false.
Your original comment:
I replied
Your response showed your usual dichomotous thinking:
This is intuitive, but utterly WRONG. Your “reject something it once accepted” is a claim that the ZnF will no longer bind to a former target. So I introduced my “Grade School” example to illustrate how the binding energies would change. Note: they ALL go UP, and the strongest binder is a new site.
You showed no indication of having read this example. Rather, you doubled down:
I grabbed my popcorn:
Only at this stage did you introduce the idea that you were referring to RELATIVE affinities (which you have yet to define…)
Your “KA_25” “refutation” is entirely anticipated in the “Grade School” example.
If you actually do mean relative affinities, then
A) you need to work on expressing yourself with at least a minimal level of clarity
and
B) your thesis is torpedoed.
Not that I expect you to understand this.
Corneel,
Exactly. If it gets in the way of cellular control like protein destruction then you probably have “bad juju”. 🙂
DNA_Jock,
Ok, so I asked:
to which DNA_Jock responded subsequently:
So at best, you didn’t answer the question question I was asking since you answered regarding a solitary finger when I was asking about TANDEM fingers, at worst it was stupid and incoherent obfuscation contrary to your claim you didn’t say something so stupid.
In any case, you lose that exchange.
But in any case, I’m thank you for keeping the thread alive.
Bill,
I’m pretty much ignoring what DNA_Jock may say from now on, I can’t get him to retract his misrperesentations, and then he answers questions I ask with answers to questions I didn’t ask. This is obfuscation and red herrings.
So feel free to ask me directly if you have questions you’d like me to address regarding any of what my critics say, otherwise some of them I just don’t have time for.
DNA_Jock, thanks for keeping my thread alive, but well, it’s simply not productive for me to keep spending time correcting how you improperly characterize my claims. So feel free to keep blasting away with stuff that I didn’t say. I’m tired of dealing with it. Thanks anyway for the conversation.
I said so here!
I went through the trouble to clarify, and that didn’t stop you from posting this drivel misrepresention after I called you out on it several times already in this thread. You just repeated what you were called out on and put it on the front page of TSZ. Oh, well, I’m not here to gain popularity points with my critics:
And no, my thesis wasn’t refuted.
stcordova,
So to get started give me the top 3 arguments why you think zinc fingers uniquely support the design inference?
stcordova,
I do think the argument about mere replication being problematic is plausible. Support would help here.
AGREED!
That’s why I appreciate your free-of-charge editorial review so I see how my words will be distorted and misrepresented and then I can try to re-write what I say so that what I say evolves more virulence against misrepresentation, red herring, non-sequiturs, obfuscation etc. So thanks for your help to that end. You’re a pal. 🙂
Before answer, this is part of a multi-part presentation tentatively entitled: “Promiscuous Protein Domains, Probability Pedagogy and Practice” to reach out to some of the IT/Engineer/Physics type people who have some biology background.
I put on the table easier alternatives to Dembski’s specified complexity arguments, show case patterns not consistent with Universal Common Descent (except by miracles), explanations for functionality of ERVs and other transposons, poly function/ poly constrained proteins, poly constrained DNA, and alternative splicing. The KRAB Zinc Finger systems and interactions are a showcase of ALL of these issues!!!
I’ll try to answer your question in a subsequent comment.
If.
And if it has a positive effect, you have good juju.
1. Some proteins have MODULAR design, and the modules appear in protein lineages that defy common descent. This is quietly acknowledged in evolutionary literature as the problem of “promiscuous domains”. ZNF136 (mentioned above) highlights the problem of trying to make genes/proteins through RANDOM copy and paste of random sections of DNA. This is analogous to writing computer code via random copy and paste!
The same problem of promiscuous domains may involve promiscuous binding motifs in regulatory regions that may regulate phylogenetically unrelated proteins. Same for miRNA motifs and PTM motifs!
2. Zinc Fingers and other binding domains and DNA-binding proteins show that DNA is poly functional — not only as a source of blueprints for proteins, but also a scaffold/parking lot/navigational aid. A case in point are enhancers on exons (WOW!) and Vitamin D receptors parking on silenced genes to regulate other genes!
There are also transchromosomal transcription factories which may leverage zinc finger proteins. Transchromosomal transcription factories/topologically associated domains are the subject of the NIH 4D Nucleome project.
3. Natural selection and waiting time problems preclude evolution of KRAB-ZFP proteins.
SO, we have a lot to talk about, and none of it is simple, but if you want to hear more, feel free to ask!
Rumraket,
What positive effect?
stcordova,
When you say promiscuous domains in comment one is that the same as what I quoted above? In this case I visualize multiple binding substrates such as cytochrome C.
Yes that is exactly the problem! A few papers acknowledge it. I know one of the researchers in the field of “promiscuous domains” personally….
Cytochrome-C isn’t the best example, the best example are KRAB-Zinc Fingers
Here is a conceptual diagram of 4 different KRAB-Zinc Finger architectures. I’ll explain more later. But for now, each line represents a particular KRAB protein architecture. The colored boxes are the domains.
ZF stands for zincfinger. Each ZF domain has spelling such as:
TGEKLYDCKECGKTFFSLKRIRRHIITH
In the protein called ZNF136 each of the 13 zinc fingers have similar spelling
SGYTPYKCKVCGKAFDYPSRFRTHERSH
TGEKPYECQECGKAFTCITSVRRHMIKH
TGDGPYKCKVCGKPFHSLSSFQVHERIH
TGEKPFKCKQCGKAFSCSPTLRIHERTH
TGEKPYECKQCGKAFSYLPSLRLHERIH
TGEKPFVCKQCGKAFRSASTFQIHERTH
TGEKPYECKECGEAFSCIPSMRRHMIKH
TGEGPYKCKVCGKPFHSLSPFRIHERTH
TGEKPYVCKHCGKAFVSSTSIRIHERTH
TGEKPYECKQCGKAFSYLNSFRTHEMIH
TGEKPFECKRCGKAFRSSSSFRLHERTH
TGQKPYHCKECGKAYSCRASFQRHMLTH
stcordova,
Got it. The promiscuity of cytochrome c was a problem for Theobald’s paper as it meant that different sequences were important to different species as electron transport was just one of several functions.
So point one is now clearer. I recommend you define promiscuity clearly in your presentation if you have not done so already.
More to follow.
stcordova,
Is your point that random mutation to improve on function can degrade another function? If so this would also apply to point 1.
Cole,
Another diagram that I put in MS Paint to show the amino acid sequences of the protein ZNF136 (metioned in the opening). There is the KRAB-A domain letters circled in orange which corresponds to the orange conceptual box to the right.
The 14 zinc fingers (1 degenerate) are also laid out to correspond to the 14 boxes on the right. Note the classical Zinc Finger architecture of a “C” letters in red with a particular spacing and then the two “H” letters in green with a particular spacing. These colored letters attach to the zinc ion.
So again this shows the concept of a domain. Zinc fingers have some obvious similarity between each other, but they can be different in spelling. Same for the KRAB-A domain.
Here is the wiki definition of a protein domain. It’s close enough for our purposes:
https://en.wikipedia.org/wiki/Protein_domain
CLICK TO ENLARGE:
http://theskepticalzone.com/wp/wp-content/uploads/2019/04/krab_znf136_domain_sequence-768×662.png
Now the KRAB domain in some proteins can be alternatively spliced out so that all that is left the protein minus the KRAB domain. This alternative isoform then has a different function. So one gene can then make two different proteins.
The full KRAB-ZFP will act as a transcription factor, but the alternatively spliced out gizmo does something with RNA!
This is an example of a polyfunctional gene/protein.
Ain’t that cool? 🙂
Pharma researchers spliced out the KRAB domain and combined it with another domain to make a powerful experimental drug for cell research. This is like plug and play for computer parts, so to speak. Promiscuous domains are like plug-and-play lego parts that show up in different proteins.
The similarity cannot be explained by DIRECT common descent. Evolutionary biologists have have admitted they don’t have an explanation. Random cut-and-paste is not a good explanation for the pattern because the cut has to be surgically precise and so does the insertion!
I’m not the only saying this is a problem, evolutionary biologists are — despite the claims by Entropy and DNA_Jock that it’s not. The literature agrees with me, not them. I prove my critics here wrong yet again!
You yourself found the paper to that effect!
stcordova,
Do you mean exons that can be alternatively spliced out?
stcordova,
This is pretty interesting in a lot of ways.
YES!
Which also means the remaining part needs an alternative promoter!
Cool beans!
Indeed.
https://en.wikipedia.org/wiki/Kr%C3%BCppel_associated_box
Plug and play lego parts! I love it.
This was the beginning of Designer Transcription Factor proteins by the Pharma companies. So we have designer man-made proteins that deliberately make a domain “promiscuous” for their own ends. The process involved surgically precise cut-and-paste, not random tandem duplication like Entropy and DNA_Jock claim for ZNF136.
God made proteins with promiscuous plug-and-play domains, and now man is learning to do the same.
Most mutations are “bad juju”, but maintaining that the regulatory network is immutable because of constraints, right after Michael Behe launched a book explaining how organisms adapt by breaking stuff is probabaly not the best timing.
stcordova,
I see you lifted the figure from the same paper I was reading. I assume that you did not only copy the pretty pictures, but also carefully read the paper.
Could you comment on the scenarios detailed in paragraphs The evolutionary path of KRAB-ZFPs and TE/KRAB-ZFP co-evolution: both an arms race and domestication, which describe an arms race with parasitic transposable elements and subsequent domestication of certain members of those? Sounds like seriously good juju to me.
I left to guys like you to highlight silly explanations.
If I described the intricacies of how a car worked and then said,
“tornadoes assembled them so that people could drive them” does that count as proof of evolution by tornadoes? Not one IOTA of analysis of the feasibility of the claim.
I started with the problem of RANDOM TANDEM duplications and then waiting time. The “arms race” claim doesn’t proceed from systematic analysis.
At one point you argued that RANDOM TANDEM duplications would inevitably end up being deleterious (“bad juju”), because every possible mutation would disrupt an existing signaling network. But if the target elements try to escape transcriptional repression (which TEs and viruses do), then mutations are absolutely required to fuel the ongoing arms race. This seemed relevant to me.
For your presentation, I have another tip for you: if any of those bigwigs you are presenting for asks you a question, then you should always try to respond to it and not casually dismiss it with some irrelevant analogy. It is a bit rude, and makes it look like you don´t have a proper answer. Are you writing this down?
What do you mean they “try”?
Jesus wants them to? So they “try”?
Are you asking because you genuinely can’t work out what he means by that?
Corneel,
You may be right with most mutations but I think it depends where they are. There is evidence that some genes have more leeway than others.
There are certainly some cellular processes that you cannot mess with to any degree or fitness is compromised.
No, there is not. There is no gene anywhere in any species that does not have any population variance, or differences between species. There is no gene that is universally conserved at a level of 100% identity anywhere across the tree of life.
Rumraket,
Where did I say 100% conserved?
Is this a new record for Bill’s sudden loss of memory?
You didn’t have to use those words, it followed logically from your claim that “There are certainly some cellular processes that you cannot mess with to any degree or fitness is compromised”. If you can’t mess with them to any degree, then mutations aren’t allowed or they would be deleterious.
Rumraket,
This is not true at all. Any degree means to degrade the function of genes that control critical functions. This would correlate to high levels of preservation not 100% preservation.