
Evolution of KRAB Zinc Finger Proteins vs. the Law of Large Numbers
There are patterns in biology that violate the law of large numbers, and thus suggests Intelligent Design or at the very least statistical miracles. The pattern involves KRAB-ZnF proteins that have multiple zinc finger domains side by side that are inexact copies of each other and would require a scenario of co-evolution of their DNA binding partners with every additional zinc-finger insertion — a scenario indistinguishable from a miracle.
The role of a zinc finger can be something like a clamp or a lock. Each zinc finger in a KRAB-ZnF protein is fine-tuned to connect with a DNA much like a lock (the zinc finger) can receive a key (like DNA). Here is a depiction of a KRAB-ZnF protein with 4 zinc fingers as part of a large chromatin modifying complex. The four zinc fingers are marked “ZN” and connect to DNA:
The evolutionary explanation of side-by-side repeated patterns of zinc fingers such as in KRAB-ZnF proteins shows a severe lack of critical thinking by evolutionary biologists who pretend “phylogenetic methods” are adequate explanations of mechanical feasibility of common descent.
To illustrate the problem, consider the KRAB-ZnF protein known as ZNF136. For reference, this is the amino acid sequence of ZNF136:
https://www.uniprot.org/uniprot/P52737.fasta
This is an amino acid fragment contained within the ZNF136 protein
TGEKLYDCKECGKTFFSLKRIRRHIITH
This short sequence is called a zinc finger domain which in 3D looks like this:
https://sciencescienceeverywhere.files.wordpress.com/2015/08/figure-11.jpg
{kind=link}
Wiki gives a description of the function of zinc fingers in proteins that have them:
http://wikipedia.org/wiki/Zinc_finger
A zinc finger domain requires two “C” amino acids and two “H” amino acids placed in the right positions. It requires a few other things too…
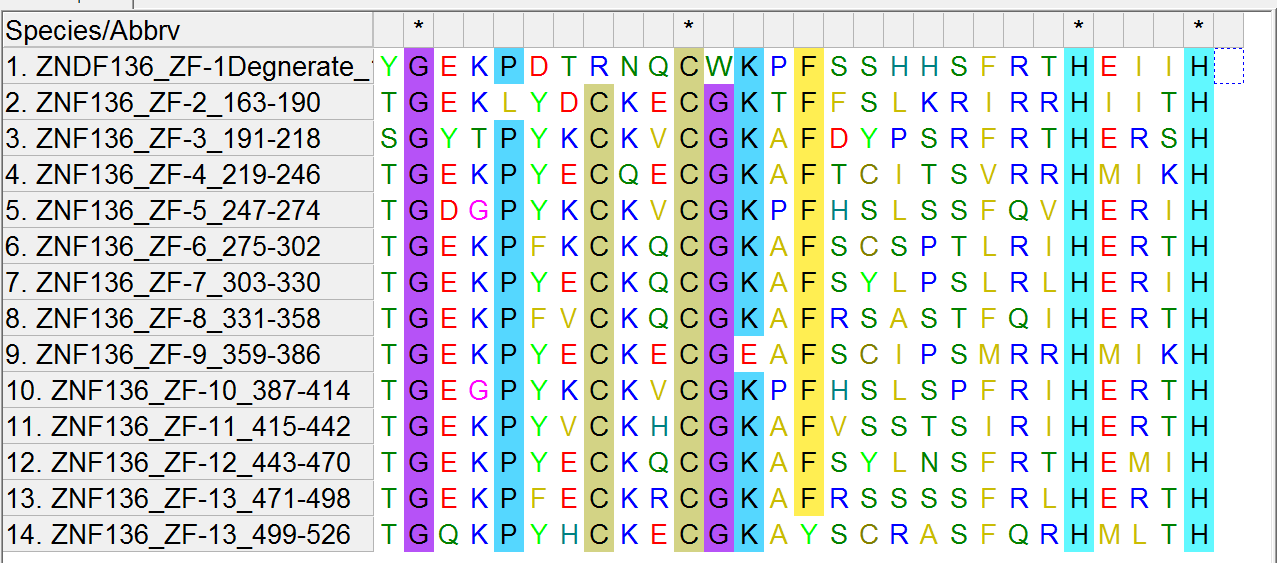
There are 13 zinc fingers in the ZNF136 protein and these are their sequences:
TGEKLYDCKECGKTFFSLKRIRRHIITH
SGYTPYKCKVCGKAFDYPSRFRTHERSH
TGEKPYECQECGKAFTCITSVRRHMIKH
TGDGPYKCKVCGKPFHSLSSFQVHERIH
TGEKPFKCKQCGKAFSCSPTLRIHERTH
TGEKPYECKQCGKAFSYLPSLRLHERIH
TGEKPFVCKQCGKAFRSASTFQIHERTH
TGEKPYECKECGEAFSCIPSMRRHMIKH
TGEGPYKCKVCGKPFHSLSPFRIHERTH
TGEKPYVCKHCGKAFVSSTSIRIHERTH
TGEKPYECKQCGKAFSYLNSFRTHEMIH
TGEKPFECKRCGKAFRSSSSFRLHERTH
TGQKPYHCKECGKAYSCRASFQRHMLTH
For completeness, there is a degenerate zinc finger in ZNF136 with the sequence “YGEKPDTRNQCWKPFSSHHSFRTHEIIH”
Why are the Zinc Fingers so different in sequence (except for the conserved amino acids)? To target a section of DNA, the zinc finger must be tuned to target it. Think of the zinc finger like a lock and DNA as a key that fits into the lock. In fact, for both the study of biology and medical applications, humans have a desire to make their own zinc fingers — like lock smiths. To bind a large segments of DNA, side-by-side zinc-fingers have to be tuned to their respective side-by-side DNA partners such as illustrated here:
By the way, there is a website that helps researchers construct the right amino acid sequence to make a zinc finger for a particular DNA target:
https://www.scripps.edu/barbas/zfdesign/zfdesignhome.php
Now, to visualize the critical/conserved amino acids, see the protein sequence here with highlights on “C” and “H” amino acids.
http://theskepticalzone.com/wp/wp-content/uploads/2019/04/znf136_zfC2H21.png
{kind=link}
Note one of the lines is not exactly like the other lines in that it is missing a “C”. This is the degenerate zinc finger mentioned above. So there is 1 degenerate zinc finger and 13 functional ones.
From this diagram it is apparent that the regular appearance of “C” and “H” is a violation of the law of large numbers, hence this pattern is not due to random point mutation alone. To “solve” this problem, Darwinist explain the pattern through segment duplication followed by some point mutation and natural selection, but this is not mechanically feasible either!
To understand why, let the reader first ponder the alignment I made of the Zinc Fingers in the ZNF136 protein using MEGA 6.0/MUSCLE software:
http://theskepticalzone.com/wp/wp-content/uploads/2019/04/znf136_zfC2H2_muscle1.png
{kind=link}
Then let the reader, consider the distance matrix generated by MEGA 6.0 which measures the number of nucleotide and percent differences between the zinc fingers.
http://www.creationevolutionuniversity.org/public_blogs/reddit/znf136_distance_matrix.xls
(All of the above results are reproducible, so I leave it to interested parties wanting to confirm the results to do so.)
For the duplication to work, at a bare minimum the right 84 nucleotide segment must be chosen, and then perfectly positioned for insertion so as not to break a pre-existing zinc finger. But supposing the duplication succeeds, why are the zinc finger’s conserved features involving “C” and “H” and other amino acids preserved and not eventually erased by point mutation given we obviously see the zinc fingers are different from each other. To preserve the “C” and “H” and other necessary amino acids in a zinc finger, the new zinc finger needs to be under selection. But in that case one is simply concocting a “just so” story for those newly minted zinc fingers without any respect for the difficulty of such a “just so” story being probable — and it is not probable!
To understand the problem of such a “just so” story, recall zinc fingers bind to DNA regions. Btw, this includes DNA regions such as ERVs! And ERVs are indicated to participate in the Stem Cell Pluripotency regulatory network:
https://www.nature.com/articles/nature13804
Further, the KRAB-ZnF protein is part of an incredibly complex machine that does chromatin modification (as shown above) by often attaching to ERV targets. But this would require that ERVs (or whatever DNA target) needs to co-evolve with the KRAB-ZnFs that attach to them!
Do these researchers even consider the fact such evolution would have to be instantaneous otherwise it would degrade function and not let the genome have the opportunity to adapt to the new accidental copy of the zinc finger because the duplication would immediately be selected against!
The above KRAB-ZnF complex is like a read/write head acting on Chromatin. Chromatin itself is an amazing mind-boggling design akin to computer ROM and RAM in one.
Again, the difficulty of evolution via random insertion/duplication mutations followed by point mutations is that such events would disrupt the binding of an already operational set of zinc fingers. For example, suppose we have an array of 10 zinc fingers side-by-side that collectively bind to a target DNA. Suppose one zinc-finger is duplicated and the number of zinc fingers is increased from 10 to 11. Oh well, the binding ability is broken or at best compromised, much like adding a single letter to a pre-existing password!
In sum, there is a violation of the law of large numbers in KRAB-ZnF proteins which is not explained by random mutation, nor random segment duplication followed by some point mutation and fixed by natural selection. Some other mechanism for the emergence of such proteins is indicated and would likely be indistinguishable from a miracle. Given the importance of such zinc finger proteins in the control of ERVs which are important in the stem cell pluripotency regulatory circuits, the origin of KRAB-ZnFs is even more miraculous.
I understand the nonsense they’re spewing, the problem is they don’t understand the nonsense.
But let’s look again at ZFP932 zinc fingers. The consensus binding was shown above and I gave reasons to show how the binding is spread to 4 zinc fingers.
Let’s say hypothetically this is the consensus binding logo of
“ACCT ”
I chose “ACCT” since it was in my illustration for ZFP932 zinc finger 1.
Ok so lets say we have the improbable event of a tandem repeat of that zinc finger in a functioning regulatory factor. Now it’s going to compromise the original binding isn’t it. The original binding was consensus:
…..xxxxxxACCTxxxxx…..
Now the new consensus binding (IF EVEN POSSIBLE) would be
xxxxxdACC?CCTxxxxx
where “?” means that it is not clear whether there is a conflict for the the fact that tandem zinc fingers target 4 bases, and when in array, there is overlap of one base per zinc finger so that there are 3 bases per zinc finger. That overlap base is therefore in conflict. But in any case, we have the situation a functioning regulatory protein is now binding to LESS targets than it did before. If this is for Transposable Elements that have to generally be repressed, this is BAD juju.
So bye bye tandem repeat. Natural selection isn’t going to let you stick around to evolve more function.
The effect of random tandem zinc finger duplication:
I see that Sal is in his triumphalist mode again, linked with his continued inability to understand the simplest evolutionary scenarios. Not a good combination.
Salvador is like a five year old who doesn’t know how to add, yet pretends to give lessons on multiplication to a professor of mathematics.
stcordova,
You’ve got several people with detailed biological knowledge telling you you are wrong. That should at least give you pause for thought.
Binding is not an all or nothing thing. Locks and keys can be a misleading analogy. If binding is strong for one part of a motif, weaker for another, you can still get an overall binding energy which is greater than that of the original motif. And, it can be tuned, in either direction, depending on the phenotypic consequences of that binding.
This is much like the amphipathic alpha helix I have discussed at length elsewhere.
If tandem repeats can’t happen, I wonder what mechanism leads to their (apparent) formation? They’re all over the place.
I won’t comment on the relevant biochemistry since that’s outside of my wheelhouse but I do find it interesting that the biologists here are talking about genuine causal forces at work in biochemistry and the critic is talking about symbol strings.
This is a frequent issue here and elsewhere. Interestingly, one of the parameters returned by a tandem repeat finder is entropy – that’s informatic entropy, not the thermodynamic quantity that increases on binding.
What you referred to as a “symbol strings” is experimentally determined consensus DNA sequences, and not very long ones at that. I provided the relevant research references of the difficult experiments involved in determining this.
The sequences mentioned in the initial post were published amino acid sequences.
Well, I’m going to present to more senior biologists on these topics in a few weeks — as in Biochem professors, Biology professors, post-docs. TSZ was my editorial review check.
Thanks to all for their determined attempt to find problems with my thesis.
One thing that came out of this is the Hierarchical Pattern that was shown by Evograd is not the result of ordinary processes of mutation and selection.
You won’t comment on the simplest of evolutionary scenarios? What, its just a ridiculously long series of molecules, which wrap together, and separate and reform, which in turn tells other molecules how to reform, and which in turn then tells cells how to divide and repeat, and change and grow at precise rates and repeat some more, and build systems, which in turn build more systems, which then form amazing series of computer like networks, which then spread over vast geographic areas, then begin to sense the geographic environments, and then sculpt those geographic environments, in ways which make the cells grow more efficiently, and which further enhances the computer networks; computer networks which can then map their own computer networks, and build some of their own synthetic computer networks, which can mimic the natural computer networks, and write songs about the computer networks, and draw paintings about the computer networks, which continue to accidentally change and make the computer network more robust, which can then copy their own computer network, and redesign the computers to fly and drive and make rocket ships to go to other geographic locations, and, and , and then have sex….all by accident.
You won’t comment on how incredibly simple this all is? Weird
John Harshman is going to be laughing at you.
So, let’s consider the RANDOM TANDEM and fixation scenario to explain even a few repeats of ZNF136.
What would the proto gene’s length be? Well without 14 zinc fingers and assuming we keep the KRAB domain and the nuclear localization signal and 1 zinc finger (all this for no good reason, but we’ll grant it for the sake of argument), oh, about 200 amino acids which corresponds to 600 bases for the coding region.
So we have 600 possible positions to start a copy, so the odds we get the right start position are 1 out of 600. The length? Nothing constrains the length to a specific number except maybe a limit of about 60 for mini-satelites, but lets grant 1-100 for range of random copy lengths.
Thus, odds of a random copy making a tandem zinc finger is 1 out of (600 * 100) = 1 out of 60,000.
What the chance of going to fixation. Well, I pointed out that if the protein is already functional as a transcription factor, the tandem repeat would in general reduce it’s binding affinity for the original Transcription Factor binding site, and worse the transcription factor starts binding to some other site in the genome! OOPS!
Not very promising for positive selection, and plenty promising for negative selection, so saying the new gene is neutral in effect is generous. We could invoke gene duplication with no expression of the copy, and the copy is where the zinc finger gets a tandem repeat. At least in this scenario the creature doesn’t have to worry about a prior function being destroyed. Probability of fixation under the generous neutral scenario?
That depends on the effective population size and structure, etc. so how about an effective population size of 10,000? Soooo, the improbability of random tandem and fixation is on the order of:
1 out of ( 60,000 * 10,000) = 600,000,000
Not exactly great odds. Recall 1-3% of proteins have zinc fingers, so this isn’t exactly an after-the-fact probability argument, it’s an argument for the probability of a convergence to a zinc-finger domain by duplication.
Then to argue selection will happen after the duplicate gene is activated for no good reason, this new transcription factor needs a target where it will participate in a new gene regulation circuit. Hmm, that means there has to be a DNA binding motif in the non-coding region of the gene it will regulate, and it has to be a positively functional regulatory change since dis-regulation is bad juju. What are the odds of that happening? Not good, but hey, don’t tell Darwinists that since they insist that such scenarios happen despite the odds (which they never calculate, but implicitly assume 100% probability) .
I’m merely questioning evolutionary premises. This is afterall TheSkepticalZone not TheGullibilityZone.
I won’t comment on the biochemistry and molecular evolution here, since that has been ably addressed by previous posters, but it strikes me that you are not being challenged on your conflation of loss of molecular function with loss of fitness. We have been bombarded the last few weeks with OPs on Michael Behe’s new “devolution” book, in which he argues that adaptive evolution will predominantly proceed by recruiting mutations that destroy or degrade function. Yet you seem to be arguing that loss and/or modification of function will be constrained by purifying selection.
So what do you think about Behe’s first law of adaptive evolution? Since the idea is incompatible with polyconstrained networks, I am guessing you don’t like it.
No Sal. The tandem finger would bind tighter and more selectively. Do you understand the chelate effect?
Your ignorance of biochemistry is impressive.
As for the rest of your flight of fantasy, what if the original Zn finger was a repressor of Pol II initiation?
How did you actually establish that? I understand that the pattern I noted doesn’t absolutely necessitate a duplication of the group of 5 ZFs, since the pattern could also be produced by selection (or design) towards a particular binding site, but I don’t see how that’s any more plausible that what I suggested.
evograd,
The only way I can think to parse Sal’s statement is that “ordinary processes of mutation and selection” is restricted to those processes that Sal believes he can model with his Hoyle-o-matic 3000.
Thus recombination is specifically excluded from “ordinary processes of mutation and selection”. See?
Sit back, relax, and enjoy the show!
What? No. Salvador thinks that everything will destroy function, that it’s impossible for tandem repeats to occur, that it’s impossible for any other repeats to occur, and that it’s impossible for selection to even happen. From the conversation it’s obvious that Salvador doesn’t even understand the simple concept of DNA complementarily, so I seriously doubt that Salvador understands what purifying selection means.
Every time we’ve tried and explained something to Salvador we’ve assumed that he had a lot more basic understanding and abilities than he actually has. When we point out that he didn’t get it, he comes back saying that he understands everything, only to then ask, or claim, something that shows that he didn’t get it at all. He’s astoundingly unreachable.
There seems to be consensus among the ID crowd that unguided natural processes are incapable of building things. As far as I know Sal believes that all mutations are destructive and deleterious. I remember that he was a big fan of the Basener & Sanford paper, which described some sort of inevitable mutational apocalyps.
So why does he bring up negative selection as an impediment to fixation? That doesn’t fit the narrative.
More selectively IF THE BINDING TARGET ACTUALLY EXISTS! If the binding target doesn’t exist, then being more selective will then reject something it once accepted.
How about fed nonsense. As in I was fed nonsense by other posters about biochemistry. But for completeness I’ll take some of the comments here before the panel I’m presenting to (which includes professors of BioChemistry and Biology) and say these are the objections:
1. Tandem repeats will increase affinity to PRE-EXISTING cis-Regulatory targets (DNA_Jock) here:
So TANDEM of the SAME IDENTICAL zinc finger, expanding it from 1 copy to 13 copies will increase its binding affinity of the Transcription Factor to the original cis-regualtory target.
2. RANDOM TANDEM is pretty much guaranteed to easily make 13 copies of of one zinc finger, but no explanation of why the correct coordinates to include a pre-existing zinc finger for copy and paste are selected. Entropy says it doesn’t matter what the copying boundaries are, he insists it’s not a problem if the repeat is ectopic.
3. Selection will select the right thing for copying or it will fix it so the repeat expansion will happen. No explanation for why selection should select for repeat expansion of the cis-regulatory sites that don’t exist or worse the tandem repeat directs the tandem repeat from the original cis regulatory site to another cis regulatory site on ANOTHER GENE. But hey, don’t let facts get in the way. Entropy and Allan Miller.
4. Some vague assertion that Sal is wrong. John Harshman.
5. The insinuation that since the 13 ZnF tandem repeats form a hierarchical pattern, and it must be a phylogeny, and therefore it must pop up naturally despite the mechanical problems already outlined. Evograd.
Well, those are the standard talking points, and I do have responsibility to make my listeners aware of objections to my claims.
#6
Oh I forgot, the one true expert here pointed out rightly the “more” tag of to create “Continue Reading” on the front page. That was the one comment, imho, that was spot on. Joe Felsenstein.
What a hopeless mangling of what I’ve said (indeed, of basic evolutionary biology).
Quite, your diligence will duly impress those professors of BioChemistry and Biology (and some assortment of post-docs too, I believe). I do recommend leaving #4 and #6 out of your bullet point list, because that would look rather silly.
Uh. My question?
So you agree that there is not much of an a priori good reason a tandem repeat of a pre-existing zinc finger should happen over any other set of coordinates in a pre-existing gene.
If you agree, then what will ensure fixation by selection of the tandem repeat. I’ve give reasons why an identical tandem repeat of zinc finger will initially degrade pre-existing function. It will be selected against on net average, not for.
The two issues:
1. probability of mutation (tandem repeat “mutation”)
2. probability of fixation
But one could argue, fortuitous point mutations happen to the tandem repeat to make it functional. Well, what’s the probability of that happening before it gets selected out or just as bad, the zinc finger no longer is a zinc finger because it got blown away by point mutation to the original tandem repeat!
Ah yes, your question. Sorry.
I heard Behe speak, didn’t read his book, so the following is my guess in relation to my claims and my understanding of Behe’s claim.
Behe says 99% of the mutations that are reproductively advantageous are function compromising.
So 99% to 99.999% of mutations are neutral to deleterious, so of the remaining 1% to 0.001% of mutations that are “beneficial” (the better term is reproductively advantageous) of those 99% or more are function compromising.
So that means a functional AND reproductively advantageous mutation happens 1%x1% to 0.001% to 0.001% of the time — like 1 in 10,000 to 1 in 10,000,000,000.
With that in mind, my point stands, the change will not be selected for.
As I have noted previously, if anyone (other than Sal) does not understand why Entropy is correct about ‘ectopic’ recombination and Sal is hopelessly confused, I will gladly try to explain.
Here’s a hint: every time Sal uses the word “copy”, he is envisioning a process of copying and pasting. Sal needs to understand cross-over.
His straw-manning of my point about binding affinities is vintage Sal, too:
Sal:
Jock:
[ N.B. For simplicity’s sake, Jock is assuming that steric exclusion is not a problem; in general, it will not be.]
Sal:
Nice try, but not what we were talking about, Sal.
Here’s an example with hypothetical binding energies — a grade school approach, if you will.
Sal’s primordial uni-finger recognizes a cognate and nine off-target sites. The energies are, in order,
5, 4, 4, 3, 3, 2, 2, 2, 1, & 1.
The DNA that codes for the finger duplicates in frame, but imprecisely.
In general there will be no steric hindrance, so ALL BINDING ENERGIES WILL INCREASE.
Now they are :
7, 5, 5, 8, 6, 3, 3, 3, 2, & 2.
The selectivity has gone up (i.e. total range is higher). The new duo-finger binds to its target quite a bit better. Excitingly, it also binds to an off-target site much better — this second site is now the best binder!
This new, avid binding could be good, bad or indifferent for the cell’s fitness.
If it is bad, it gets selected out. Either the second site evolves, the duo-finger evolves, or the duo-finger goes extinct.
If it is indifferent, drift at the second site.
If it is good, then novel effing function just arose!
In all cases except the ‘goes extinct’ scenario, you have a new and improved Zinc finger protein.
I am embarrassed for Sal.
DNA_Jock,
That was a beautiful explanation. Wasted on Salvador, but excellent explanation nonetheless.
Just so I can look so official I’ll do my imitation of being evolutionarily saavy.
I made the following phylogentic tree of the ZNF136 zinc fingers under these parameters to showcase irrelevant mathematical theatrics to at least suggest I found some profound evidence of common descent and natural mechanism of evolution as an explanation of the patterns of the 14 zinc fingers. I even inferred the evolutionary history! The auto generated caption said so:
By golly this proves it happened naturally. My software run shows “the evolutionary history” Dab Gummit.
It would be a shame if all this hard work was lost. With a little bit of effort you could create something that’d last. For example if you were to prepare the figures and tables, write the methods, write up the results, write the discussion then finalize the results and discussion before writing the introduction and then write a clear conclusion and a compelling introduction plus an abstract then compose a concise and descriptive title and of course select keywords for indexing and finally write the acknowledgements and references you’d have something you could potentially put your name to and publish!
You already have most of it right here already. Why not submit a real scientific paper to an ID friendly journal?
https://www.elsevier.com/connect/11-steps-to-structuring-a-science-paper-editors-will-take-seriously
Now if the phylogeny in the above diagram (or any phylogeny for that matter) was constructed, and even assuming the phylogeny was correct, it does NOT solve the following problems:
1. Random Tandem with zinc finger multiplicity of 13-14, and side by side to boot! A phylogenetic diagram doesn’t explain the close proximity of the random tandems on the same gene. If these were dispersed repeats, then there would be a problem with random tandems, but they tandems are there.
2. If these were in non-coding regions and we didn’t know about the binding specificity of zinc fingers, then maybe the phylogenetic interpretation would hold, but we know this protein is a transcription factor with a cis-regulatory target, and that zinc fingers are relatively specific in sequence since their cis-regulatory targets are specific.
The zinc fingers are not subject to random evolution without compromising function. One could of course invoke the absurd scenario that has been repeated in literature that the cis-regulatory element changes to match the zinc finger and vice versa. The problem is there are MULTIPLE locations of the same cis-regulatory motif as the same Transposable Element appears in the regulatory region of multiple genes! So one has to invoke the absurd scenario of multiple copies of cis-regulatory elements in mutiple genes simultaneously evolving to match the zinc finger. That’s a statistical miracle to have copies of the transposons conspire to have the same point mutations!
Ok, we’ll use John Harshman’s theory that common descent is compatible with miracles.
But what does this show. NESTED HIEARCHY is no proof that common descent proceeds naturally. ZNF136 is case in point.
And if we can refuted common descent as an explanation of NESTED HIERACHY in one case, we might be able to do the same for common descent in the rest of a proteome. And if we can do that, we might be able to refute common descent as an explanation of nested hierarchy across species. If so, common design is the best explanation.
Sal,
This may be a teaching moment. Could you describe, in your own words, what point evograd was attempting to get across with his phylogeny diagram, and precisely what aspect of the diagram supports his contention?
The evolutionary history of what, exactly?
But as an alternative it seems you are proposing a literal miracle instead?
Evograd was showing a nested hiearchy. I did the same thing.
Evograd rooted his phylogeny with the degenerate zinc finger, I made an unrotoed tree.
You can ask him what he was trying to do, but it looks like he was suggesting a way a duplication scenario would happen.
But, you can see my unrooted, unprejudiced tree shows the sequence nearness and it does NOT agree with his tree.
The rooting of the tree by Evograd was not justified by anything unless there is bias making tandem repeats from 5′ to 3′ directions only (relative to the direction and sense strand the gene is read from.)
One could of course argue the ZNF136 came from similar zfp that looks like ZFP932 which has only 4 tandem repeats. But we can likely rule out ZPF932 specifically since it is mouse specific, but something like it might work.
Of course, one has just shifted the problem away to another gene, but the cumulative problem of random tandem over many generations of genes remains. It doesn’t solve the problem.
What does then? Any hints?
What a lovely idea.
That’s a long chain of if, each if of decreasing likelihood. You end up with likelihood so low that you might as well not have brought it up. You haven’t even managed the first if.
About 1-3% of proteins in C. Elegans have zinc fingers. If similar proportions hold for humans, then about 200-600 proteins have zinc fingers.
The architectures of proteins with zinc fingers vary. You can see the location of them in a variety of protein families at the NIH CDART database:
https://www.ncbi.nlm.nih.gov/Structure/lexington/lexington.cgi?cmd=cdd&uid=338531
Here is a screen capture of only 1 page out of the 24 showing known protein families with predicted zinc fingers. There are likely more families with zinc fingers that didn’t appear in my CDART search.
Clearly, in light of such CDART diagrams, there is no requirement for random tandem to make zinc fingers line up side by side so nicely as in ZNF136.
The reason it is functionally desirable for tandem zinc fingers to appear in a protein is that a solitary zinc finger can only specify a DNA binding target of at most 4 bases, which is not much specificity. ZFP932 (discussed in the comments) has a cis-regulatory target of 12 bases where there there is an overlapping base that is bound by adjacent zinc fingers, thus we have 3 bases targeted per zinc finger even though a solitary zinc finger can target 4 bases.
CLICK TO ENLARGE:
http://theskepticalzone.com/wp/wp-content/uploads/2019/04/zinc_finger_cdart-1024×453.png
stcordova,
If you ever get to talk to a biologist, show them evograd’s diagram and comment, and your response.
Your inability to read for comprehension will amaze them.
I really cannot think how evograd could have made his point (and the nature of his supporting evidence) any clearer and yet you missed it. Spectacularly.
DNA_Jock,
Can you elaborate why you think Evograd’s argument was not comprehended?
How is this relevant to whether there is evidence for an evolutionary path for these proteins or is this just an attempt to slip in an ad hominem?
Are you still a moderator?
I decided not to read Salvador’s comments any more, since he has missed each and every one of my explanations, spectacularly. I thus predicted he would not understand evograd’s example either (even though it’s actually simple, it requires more understanding than the even simpler expansion of repeats because of DNA complementarity). So, whatever DNA_jock might be pointing out, I don’t even need to see it to be sure that my prediction was fulfilled.
Did you see evograd’s comment? If you did, you’d see that it shows data compatible with one section being a copy of the other section, thus providing an evolutionary path. Combined with my explanation about tandem duplications, it should be very easy to put two and two together and understand how that’s evidence of an evolutionary path. If you saw it but did not understand it, then you should say so, that you didn’t understand it, rather than advance the hypothesis of an ad hominem.
Remember, ad hominem is attacking the person over the argument (the argument is rejected because the person is stupid, but the argument should be evaluated on its own merits, not on those of the person presenting it). Insulting the person because of the stupidity of the person’s argument is not ad hominem, it’s a conclusion (the person is stupid because he/she presents stupid after stupid arguments, the person is unable to notice the stupidity of the arguments, and never learns from those mistakes).
Wha? I’m struggling to parse that. I don’t know what you mean by coordinates. Tandem repeats are a form of mutation. They happen at some frequency. Some will be selected against, some not. We would expect to see a preponderance of those not-selected-against in surviving genomes. Does that seem wrong to you?
I don’t see how degradation of existing function is universally to be expected, at all.
Some mutations are detrimental, therefore all are, you seem to be saying.
I believe that you were trying to argue that duplication of the zinc finger was compromising function, so I don’t think you should be multiplying with the chance that it is still functional.
Anyway, that is besides the point, because that wasn’t what I was asking. Trying one final time: What do you think about Behe’s first law of adaptive evolution? Since the idea is incompatible with polyconstrained networks, I am guessing you don’t like it.
Just trying to get some idea about your position.
Entropy,
Jock attacked Salvadors competence and not his argument. Thats against site rules and he is moderator. People are wondering why this site is going down hill.
Single evidence of a duplication is a long way from demonstrating a Darwinian explanation for zip fingers. At this point both sides are pivoting and talking over each other.
Apparently my comment needs some clarification. My original comment and figures had nothing to do with the nested hierarchy pattern. The point I was making with the phylogeny was simply that ZF #4 clustered with #9, #5 with #10, etc. This suggested to me that ZFs #4-8 duplicated to give rise to ZFs #9-13.
I didn’t say anything about the deeper nodes in the tree for a reason. I’m wary of trying to reconstruct a meaningful phylogeny with such short sequences. I just noticed the rough pattern that seemed to suggest a duplication event and decided to mention it for the benefit of the discussion.
With pleasure.
Evograd noticed (by eye, no less!) that repeats 4 through 8 bore an uncanny resemblance to repeats 9 through 13, almost as if the repeat number had jumped from 9 to 14 via a single cross-over event, as in
ABCDEFGHI becomes ABCDEFGHDEFGHI
I hope you understand that a single homology-driven crossover can produce this result, because Sal evidently does not. Just start singing verse D instead of verse I.
So evograd made the repeats into a tree.
The dispositive thing about this tree is that repeat #4 is most closely related to #9, 5 and 10, 6 and 11, 7 and 12 and 8 and 13. Which is exactly what you would expect if they had arisen via a (relatively recent) single
ABCDEFGHI becomes ABCDEFGHDEFGHI
event.
It’s fricking amazing! It is vanishingly unlikely under Sal’s Hoyle-o-matic view of cutting and pasting.
The question is “Does Sal understand this?” [As an aside, the question is not “Is Sal right?”… When he disagrees with a reality-based commenter here, Sal is yet to be right about anything.]
So I ask him to paraphrase evograd’s point — let’s see if he understands what he disagrees with.
Given the opportunity to demonstrate his comprehension, Sal whines an irrelevance about the rooting of the tree (perhaps failing to notice that “degenerate” repeat 1 isn’t even a full Zinc finger) and waffles some utter rubbish about
Proving that he cannot get past his Hoyle-o-matic thinking.
He utterly failed to pick up on the significance of the 5-9, 6-10, 7-11, 8-13 pattern.
It’s horrendous.
I believe that the tree-drawing program he is mis-using assumes that all 14 sequences arose via bifurcation, but I will let someone with phylogenetics expertise weigh in on the dangers of dropping sequences into automatic tree-drawing programs that you don’t understand, complete with auto-generated captions!
I really am embarrassed for him, and I am still a moderator.
I’ve been ninja’d, but I am going to post this anyway, without reading the intervening comments. Let’s see how we all do…
False. Jock pointed out that Salvador’s incompetence was on display yet again because Salvador didn’t understand evograd’s comment and figure.
This is not about whether there’s a Darwinian explanation. We’re discussing the natural mechanisms that give rise to observed patterns. The role of evolution (not just Darwinian), is in the selection of resulting proteins that have some function, and in keeping important positions in the sequence from diverging. But Salvador doesn’t even understand the mechanisms. He’s too incompetent to understand them as he has demonstrated consistently, not just in this thread, but also in previous ones, where he even quoted from articles that contradicted his intended “points.”
Salvador is like a kid who neither knows, nor understand, addition, yet pretends to lecture math professors on multiplication. The professors have assumed that the kid knew how to add, explaining multiplication in many different ways, but now they’re amazed to notice that addition is way above the kid’s level. Evograd’s comment is like trying to explain exponents to the same kid. Of course Salvador won’t get it. He doesn’t get addition in the first place.
This is why I stoped. I don’t know how to reach a kid who doesn’t understand the very concept of addition. The kid keeps misquoting from wikipedia hoping to hide his incompetence that way, but the misquoting only shows, much more evidently, that addition itself is beyond his reach.
DNA_Jock,
Why don’t you and Entropy start by demonstrating you understand Sal’s arguments. You guys are talking over each other. You as a moderator need to be a leader and right now you are leading this site right off a cliff.
DNA_Jock,
Does anyone have experimental evidence that these sequences have flexibility? Are they highly preserved?
Yep. Pretty sure the rooting is actually irrelevant. I’d bet if you plug in all seqeunces, don’t define a root, you’d still get the same relationships with (5,9)(6,10)(7,11)(8,13).