
Dr. Winston Ewert put forward his module hypothesis, but I put forward an alternate module hypothesis at the domain and motif level of proteins. It is based actually on papers by evolutionists who have pointed out that the problem of “Promiscuous Domains” remains an unsolved problem in evolutionary biology.
When I put Promiscuous Domains on the table in the Common Design vs. Common Descent thread, the TSZ Darwinists ignored the problem and then declared victory. I viewed their non-response as evidence they didn’t understand the problem and/or preferred to ignore it.
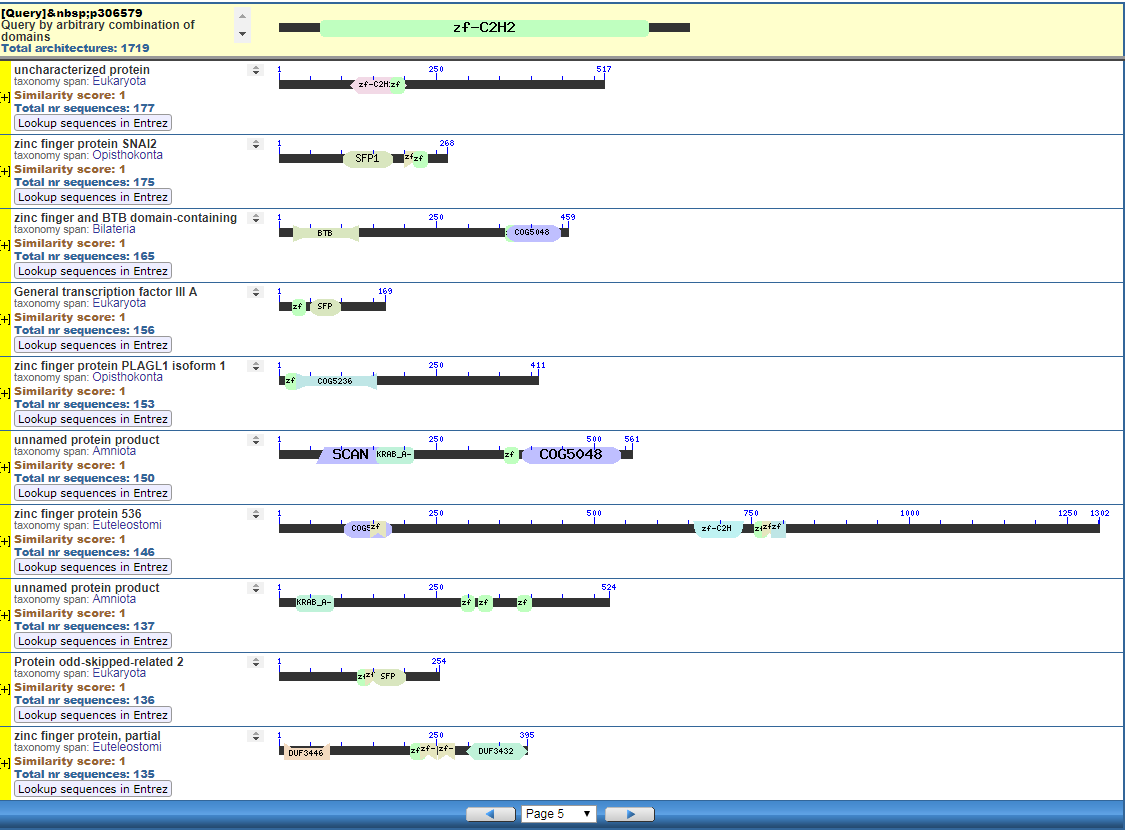
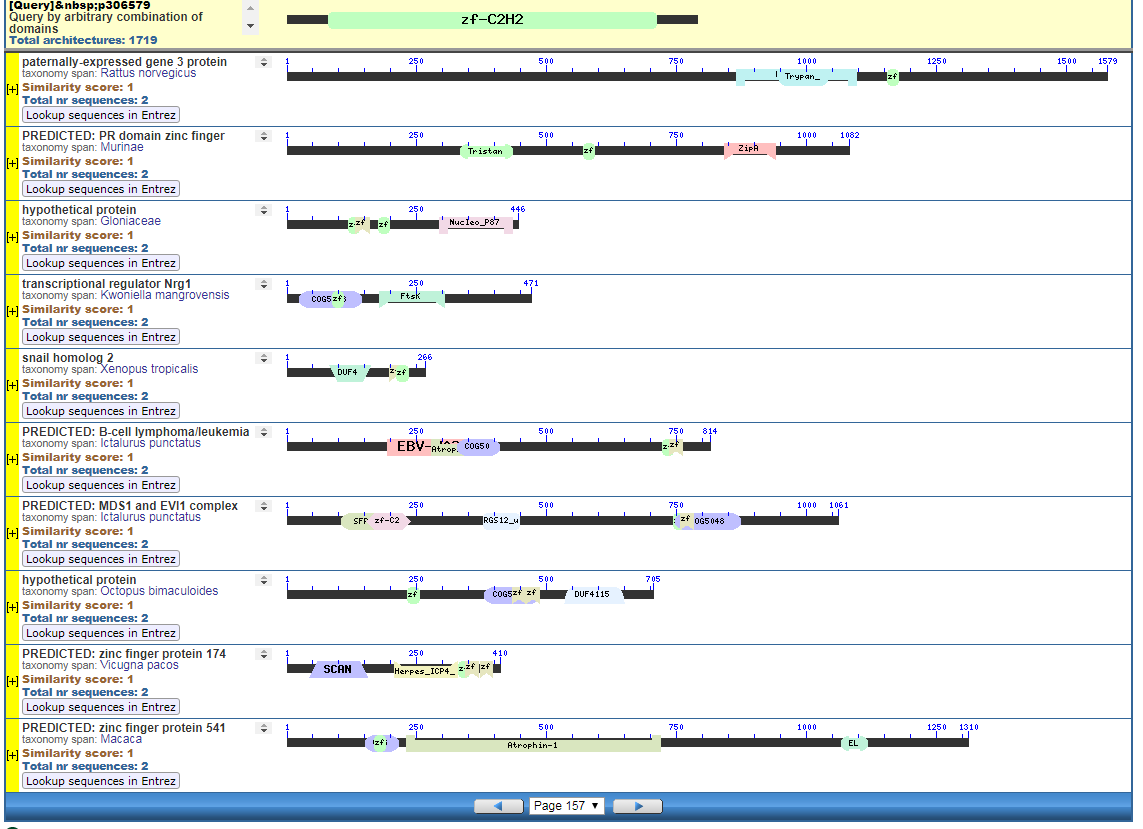
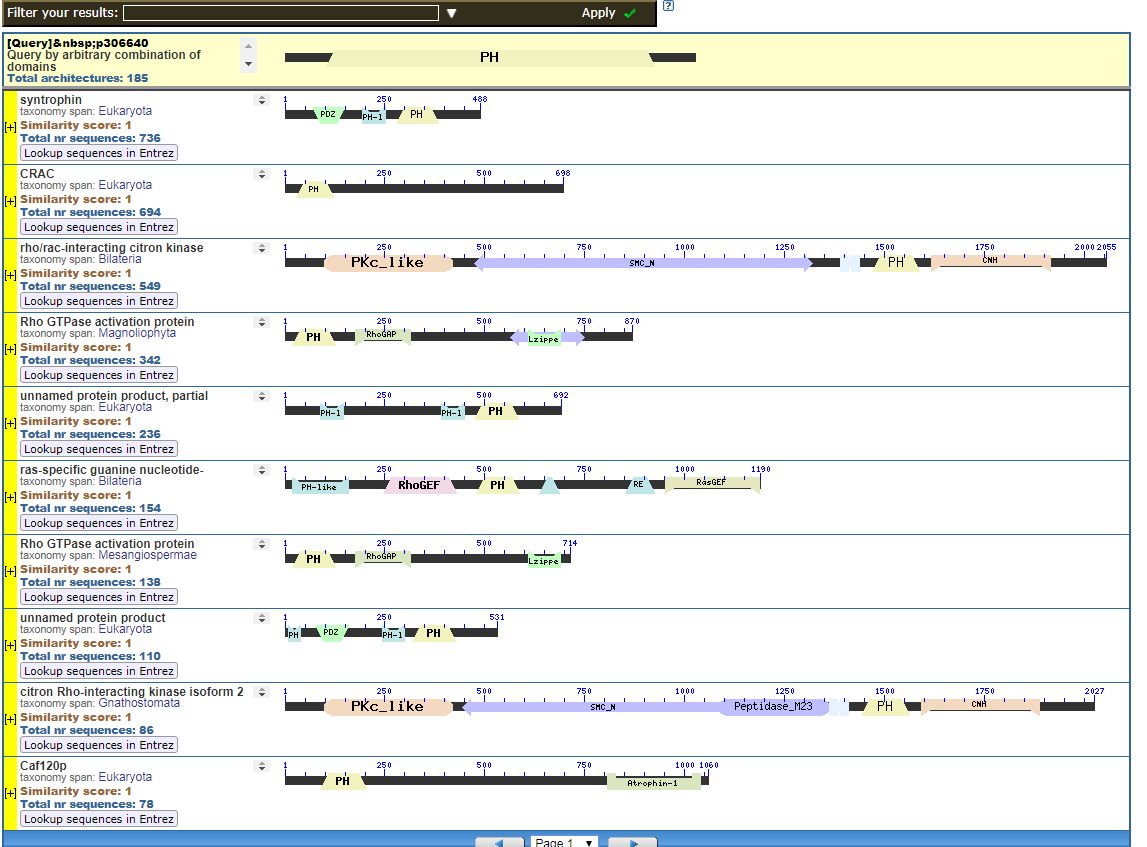
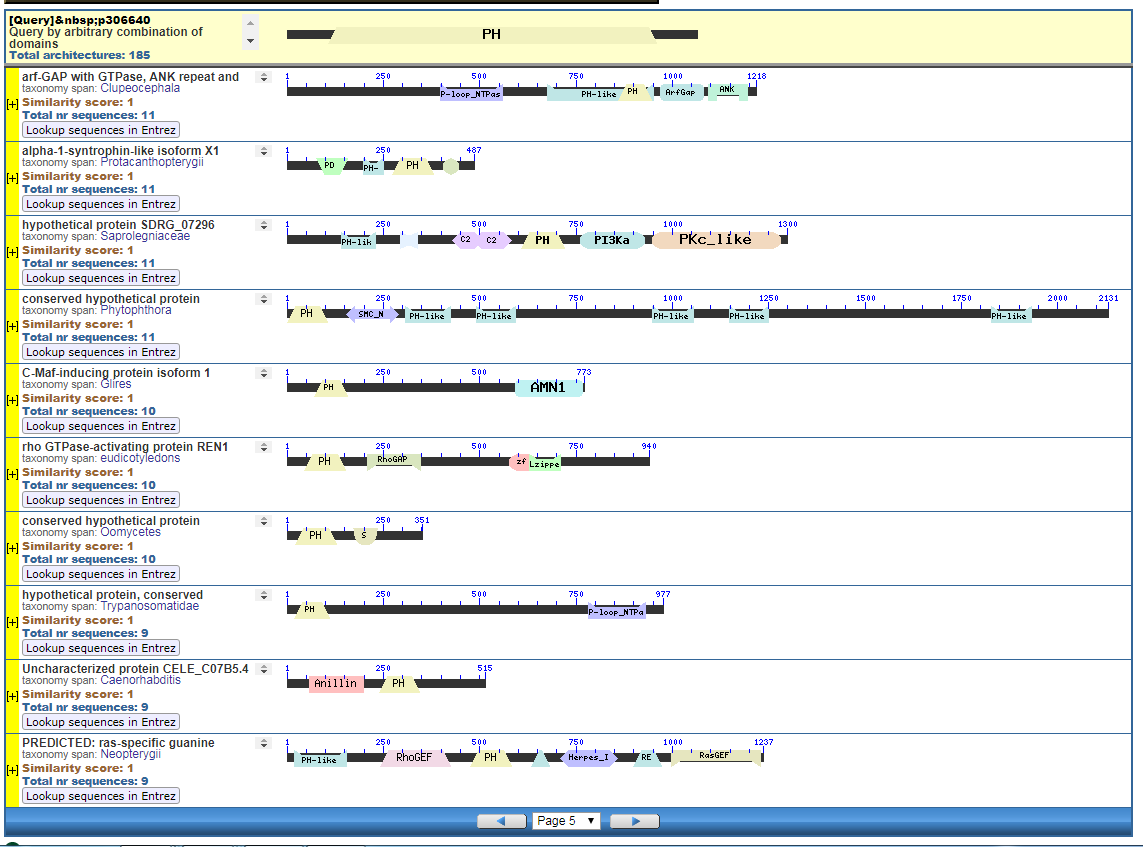
Perhaps pictures are worth ten thousand words. From the NIH, that great source inspiration for the Intelligent Design community, we have the CDART database viewer.
From the CDART viewer, I provide a few of the thousands of diagrams that show the promiscuity of protein domains. The diagrams below show the classical zinc finger ZF-C2H2 “ZF” domain and the Plextrin Homology “PH” domains. Note how the location of domains is “shuffled” to different locations in different proteins. It’s as if proteins are made by different lego-like parts in different order and position. My preliminary look into small 4-amino acid motifs that are the target of phosphorylating kinases suggests the the problem of promiscuity goes all the way down to small motif levels.
Such promiscuity is more consistent with common design than common descent.
Click to Enlarge Classical ZF-C2H2 Zinc Finger Page 5
Click to Enlarge Classical ZF-C2H2 Zinc Finger Page 157
Click to see all CDART Classical ZF-C2H2 Zinc Finger Architectures
That’s right.
That’s wrong. The point of the graph is to point out the sequence differences, right down to even 4-amino acid motifs, HECK, even a single amino acid (like a Serine or Threonine) in the right location makes a difference. That’s not consistent with random mutation in the process of common descent, at best it is consistent with little acts of special creation with common descent, but if one starts invoking little acts of special creation, then what’s the use of common descent, the hierarchy is just as consistent with special creation by a Designer who delighted to build things with hierarchical relationships to facilitate scientific discovery.
I demonstrated this with the critical metal binding proteins and two phosphorylation sites (one in Top2A and one in Top2B). There will be more to come.
Bottom line: either you think the patterns of similarity and diversity are the products of mutational accidents, or they are the products of a design. I’ve provided evidence that even small motifs cannot be accidental because it would compromise function.
Same false dichotomy. Either a complete single-point-mutation accident or god-did-it. Always neglecting to think of recombinations, which are also naturally occurring mutations of a larger scale than point mutations, and the effects of negative and positive selection in filtering out the results of those “accidents.”
Plenty of sensical options instead of going all the way coo-koo with that fantasy about magical designers.
You provided evidence that you cannot look into the appropriate literature, that your ability to verify domains, or motifs, doesn’t go beyond presenting pretty pictures, that you don’t know what you’re talking about, that you cannot understand the most basic concepts, and that you can only read for quote-mining.
No it’s a demonstrable fact. All your silly little declarations you end your posts with are in no way substantiated by listing the random facts you do.
That’s a textbook non-sequitur right there. It simply doesn’t follow.
If you disagree, please try to put it into proper syllogistic form so we can see how you derive your conclusion from premises.
It would have to look something like this.
Premise1: Topoisomerase Top2A interacts with… ?
Premise2: Proteins X, Y, and Z have these motifs … ?
Premise3: … ?
Conclusion: Therefore it is not consistent with random mutation in the process of common descent.
What is missing is the actual connection between the facts and the conclusion you mindlessly declare.
Look, any idiot can do what you’re doing but in reverse, and it will look no less mysterious:
The point of the graph is to point out the sequence differences, right down to even 4-amino acid motifs, HECK, even a single amino acid (like a Serine or Threonine) in the right location makes a difference. That’s completely consistent with random mutation in the process of common descent, but not with special creation.
You didn’t demonstrate shit. You seem to be laboring under the delusion that merely declaring something constitutes “demonstrating” it.
Compromise function of what? And why would the emergence of a new motif compromise function?
Where did you provide this evidence? All you did was list the presence and types of some motifs. In fact what you’re saying makes zero logical sense.
The general idea with any new beneficial mutation, whether emergence of a new motif or not, is that before that mutation happens it isn’t strictly necessary to the survival and reproduction of the organism. But once it happens, it has a potential to positively affect the fitness of the offspring that gets that mutation, and can then later in conjunction with other mutations(or environmental selective pressures changing), become critically important for function.
The same would apply for the evolution of a particular motif, such as a phosphorylation site in a protein (say). Before a particular motif evovles, it’s putative future function isn’t needed, at best merely beneficial when the mutation(s) whereby the motif comes to exist in a particular protein, happen. Subsequently, this emerged and beneficial motif becomes increasingly important so that it later becomes critical for function. In effect a version of this is exactly what happened in the LTEE. The evolution of the cit+ function under aerobic conditions is conditionally beneficial, and can even be thought of as conditionally critical to survival, if the organism switches to an environment with the only carbon source being citrate, then it is absolutely essential. And just like with a motif being transposed or duplicated into a particular protein, a duplication or transposition of an already existing protein coding gene into another area of the genome which is immediately downstream of a promoter active under aerobic conditions is, in essence, the same genetic phenomenon(a piece of the genome from one place, is transposed or duplicated into another place, where it turns out to be beneficial).
Saying that this can’t happen because it would “compromise function” makes zero logical sense. Compromise function of what?
If the fossil record is young and life on Earth is young, the nested hierarchical pattern is not explain by common descent. The Designer thus had his purposes for the nested hierarchical pattern.
We don’t need to know all the reasons for the pattern, but if it is there and is useful for predicting chemical activity, that is sufficient to leverage the patterns for biotechnology and medical benefit.
If life on Earth is old, the argument against common descent is harder, however if shuffled domains and motifs are real and if they are chemically significant, then this is convergence, and hence the product of comomon design, not common descent.
Rumraket,
So how do you explain 3% of the human proteins having zinc fingers. Do you think that was the result of gene duplication or convergence?
Why?
Three false premises. The earth is not young and life is not young, and you can believe that only by ignoring all of geology and physics. Third, even if life were young, there would be no other sensible explanation for nested hierarchy other than common descent. It would require vastly speeded up evolution, certainly forced by the creator. That makes no sense, but it makes better sense than a pointless created hierarchy.
Ignorance is strength? The fact that we can use the pattern to our benefit is not a reason to suppose that it was created for that reason, right?
The argument against common descent is hard enough that you seem unwilling even to try to make it. Anyway, convergence isn’t evidence for common design, much less proof. I will note that once again you are using “common design” very ambiguously; once you supposed it to be a synonym for separate creation of species, and now apparently just for creation of motifs. Any meaning it may have had is leaking away rapidly.
First of all I think the overwhelming majority evolved before the human lineage split from our common ancestor with the chimp and probably over the entire course of eukaryotic evolution, and probably extends back before this as DNA binding proteins is not a uniquely eukaryotic, much less vertebrate, mammalian or even human phenomenon.
I think most DNA binding proteins (transcription factors and zinc fingers among them) evolved by duplication and divergence.
Sal, I some times suspect that you come here just because you recently took some (probably entirely interesting) course in molecular or cell biology, but then you feel compelled to try to square your latest gained knowledge with creationistic apologetics.
Whether there is even the slightest merit to attempting such an endeavour seems to be of no importance to you. You learned something new, and then you want to show it off while evangelizing for christian creationism.
Rumraket,
I think Sal has been quite open about that.
I keep coming back to see if there’s anything new on the ID front. What I see are variations on paley, updated with the latest technobabble. I appreciate the patience with which some of you dissect this corpse.
But until the creationists actually come up with ideas for research, they are on ignore. Sometime I see something interesting and decloak them, but not often.
Saves a lot of wear and tear on the brain.
With some difficulty, I created the following diagram (below) mostly from scratch with the help of MEGA/MUSCLE and PhosphoSitePlus data and MS Paint. The very first row is human Top2A protein and the very last row is human Top2B protein with other creatures in the rows in between.
The arrows point out residues that have had experimental reports of post-translational modifications in Humans.
I personally believe it’s entirely possible many other positions have not yet been reported as yet for post translational modifications since many post translational modifications (PTMs) are likely cell-phase and cell-type and developmental-phase dependent!
The text connected to the arrows is hopefully an understandable short hand. For example:
S1525p = serine at position 1525 in Top2A is phosphorylated
K1492ub = lysine at position 1492 in Top2A is ubiquinated
K1492sm = lysine at position 1492 in Top2A is also sumoylated
K1480ac = lysine at position 1480 in Top2A is acetylated
etc.
This also shows functional optimization for each species. Other creatures cannot in principle have the same set of PTMs as humans since they don’t have the same amino acid sequences!
Where this will lead possibly, is that for these post-translational modifications to happen, an addressing scheme needs to be in place for the proper machinery to locate the protein and the short amino acid segment (say 4 amino acids) in a sea of other proteins. So not only is there species specific post-translational modification, but species-specific machines and addressing systems to make this possible.
I believe the Designer has optimized the patterns of diversity to help us figure out the small motifs (say 4-amino acids) that are used to co-regulate sets of proteins. For example, the motif “rkps” is found in numerous proteins and I believe it is the target of a machine called CSNK2A1 which locates proteins with the address of “rkps” and phosphorylates them. That is what is implicitly suggested in the PhosphositePlus website where I got some of the data to construct the graph. I simply enter “rkps” in the PhosphositePlus website and it lists the proteins that are phosphorylated with that 4-amino acid motif. This suggest in addition to “promiscuous” domains, there are also promiscuous motifs.
The graph also strongly argues that there are indeed functional differences in the paralogs of Top2A and Top2B which are created from separate chromosomes in humans.
Given PTMs result in a metabolic load, it is hard to believe creatures expend so much effort to make these species-specific PTMs for no good reason.
Click to ENLARGE
Continuing from the previous comment, here is the next adjacent segment on Top2A and Top2B. Again, the PTMs are obviously different on the parlogs, not to mention, in principle must be different on the other species. Note some of the specialized PTMs on Top2A like the 1st degree methylation on Argenine (R1443m1).
John Harshman, Rumraket, and my other detractors basically ignored the problem of the phosphoproteome, the acetyl proteome, the methyl proteome, and others like the sumoylation “-ome” the ubiquitination “-ome”. John Harshman often accuses me of making up “-omes” until I call him on it by showing most of these “-omes” are acknolwedged in one way or another in the literature. These diagrams I’m providing hopefully illustrate these “-omes” exist, and there is no small amount of machinery to make them happen!
Click to ENLARGE
Extending the previous comment, here is the next adjacent segment.
Click to ENLARGE
What you believe is uninteresting until you can present some kind of evidence and argument in favor of your belief. Why would such optimization result in a nested hierarchy, much less the same nested hierarchy for different loci?
It doesn’t have to.
You’re obsessed with explaining the nested hierarchy as far as the designer’s reasoning. That doesn’t have to be explained as much as Mount Rushmore doesn’t have to be explained. The issue is whether the organisms are the expected outcome of natural processes. If they are not the result of natural processes, then they can be reasonably argued to be products of special creation, and if products of special creation, the nested hierarchical is also specially created.
The evidence that the hierarchy highlights functional features has already been confirmed at the level of metal connecting residues or catalytic residues conserved across all species in a hierarchy. The conserved residues in the sub portions of the hierarchy which nest in a larger hierarchy should highlight lesser functions. That is a testable prediction.
By your criteria, not mine.
The next adjacent segment I just completed and I present it below.
Click to ENLARGE
excellent point Sal. I think this is the core of the misunderstanding among critics.
All “our side” is trying to demonstrate is that X is not what you’d expect from natural processes…….Full stop
Unfortunately “the critics” seem to be having a different conversation, one about what God would or would not do.
peace
False, straightforwardly false. This thread is CHOCK FULL of declarations from Sal that what we see is what the designer would design and only makes sense on design.
And this is where we show, analogously to what you’re trying to do with natural processes, that what we see is not what you’d expect from an intelligent designer.
It seems to me we can have both conversations. What would we expect from a natural process, and what would we expect from an intelligent designer, and then we can compare those expecations to what we actually see.
What we see is what we would expect from a natural process (nesting hierarchical structure in the data, consilience of independent phylogenies), and it is NOT what we would expect from an intelligent designer, because an intelligent designer has no good RATIONAL reason to design nesting hiearchical structure in the data, much less consilience of independent phylogenies.
Thank you and have a nice day.
Sal in the op:
“Such promiscuity is more consistent with common design than common descent.”
Sal in a post responding to John Harshman:
“To optimize scientific discovery. Did you ever look at alignments of zf-C2H2 alignments. It’s pretty obvious what the critical residues would be in a 3D conformation. That would not be evident if the zf-C2H2 fingers were identical.
It would seem the designer had us and our capabilities to comprehend in mind.”
Another post:
“At ICC 2018, I talked with my colleagues and insisted if life is young, then the hierarchical pattern is by design. The question is why? My answer: God wanted to make it easier to decipher biological function.”
Another post:
“It’s more subtle than that, what if the Designer wanted to highlight multiple functions, he could do so with a hierarchical arrangement. Random variation will not work, btw, for co-regulated motifs spread across multiple proteins, as I was trying to show with the PhosphoSite motif search.”
Another post:
“A reasonable explanation for the spread of zinc fingers of many varieties over many proteins implementing coherent function is common design, not common descent:”
At which point it looks like Sal realized he can’t support any of these conjectures, so changed his tune to just declaring brainlessly that the distributions of various domains or motifs “don’t look random” and therefore aren’t “explained by accidents”.
A designer, not necessarily God.
Nobody has ignored anything. We are still missing a coherent explanation of what the problem is even supposed to be.
You declare it is a problem, but never get around to show in what way it is a problem. Much less do you show how you intend to solve it with a superior and testable scientific hypothesis.
Basically your whole case here consists of pointing to some random factoids (with lots of pictures and technobabble that is sure to impress someone who doesn’t have a clue) and saying, without any explanation of how or why, that it isn’t consistent with common descent, but instead is with “common design” (which you never define or model in any way).
Depending on the audience that is a pretty good design.
Not a reasonable response.
But Mount Rushmore does have to be explained. You aren’t making any sense here.
None of that follows. If (and it’s a big if) certain features of organisms have to be explained by special creation, then special creation of just those features is the most parsimonious explanation, and other features can have their own most parsimonious explanations. The nested hierarchy is most parsimoniously explained by common descent. Put that all together and you have guided evolution, not separate creation of species. The only reason you reject that hypothesis is your prior preference for separate creation, and that preference has nothing to do with science or with anything empirical.
I don’t think it actually is. And it doesn’t explain why the pattern is a nested hierarchy rather than any other pattern, nor does it explain why different loci show the same nested hierarchy.
What did that mean? I don’t think you can defend any of your claims. You’re just saying the first thing you think of and then refuse to examine its implications.
Rumraket,
Technobabble Rumraket? I showed species specific differences in amino acid sequences on the same protein across species have species specific function, namely the post translational modifications. Random accidents don’t make function any more than tornados passing through a junkyard make 747’s.
John,
If biological organisms are specially created, that is a sufficient, not but not necessary explanation to refute common descent.
Yeah that’s the technobabble part. It’s technobabble because it doesn’t accomplish anything other than it seems to be present only to give a superficial appearance that your following claims have some sort of technical merit.
I agree, but then again tornadoes, junkyards, or Jumbo Jets have nothing to do with the process of biological evolution. So this particular conclusion is just irrelevant.
What you mean to say is something like this:
[A collection of technical facts] -> [Therefore X is inconsistent with evolution]
The problem is that your [collection of technical facts] does nothing to substantiate your conclusion that [X is inconsistent with evolution].
When you repeat this same silly method over and over again, without ever tying your technical facts together with an actual model of evolution to show that the random facts are inconsistent with the model, that is what makes it obvious that your collection of technical facts is only there to give your unjustified conclusion the superficial appearance of having some technical basis in fact.
But the connection is missing every time. There’s the facts, and then some stated conclusion that never follows. So your facts are just there as impressive filler. Lots of abbreviations, lists, graphs, and figures. And then some COMPLETELY out of the blue conclusion, usually represented in the form of some deliberately sill caricature of evolution (like the tornado in a junkyard, which has NOTHING to do with how evolution works).
Sorry, the gig is up. Your behavior is more transparent than intergalactic voids are to the passing of neutrinoes.
And what a big if. The problem is that the evidence doens’t support that claim. In fact the evidence points to common descent, so one wonders why you would believe organisms were specially created since nothing in biology supports that conjecture.
It isn’t even clear what would actually follow from special creation if we don’t allow ourselves to actually speculate on what the designer was trying to accomplish or what process of design and manufacture were used. If we refrain from speculating on such matters, then we can’t have a special creation model that makes testable predictions, and thus no observed pattern could be said to be evidence of the model (since we don’t have a model).
I have on occasion suggested that I don’t think the Christian God would deliberately design objects with false apperances of age, distance, and genealogical relationships. That’s just me I guess, you seem to be of a different persuasion and to you it is entirely reasonable that God would go out of His way to construct completely deceptive patterns expected of common descent for no good functional reason. I admit outright that I have no way of proving that a God could not do such a thing. But I also have no good reason to actually believe God would do that.
So since the evidence before us nevertheless IS the kind of evidence that common descent is expected to leave behind, I’m going to suggest it is most rational to interpret that evidence as being the product of common descent, rather than the product of God going around planting evidence to frame a process that didn’t actually take place.
You believe what you believe, and you give the interpretations of the evidence that you do, because you seem to have prior commitments outside of the actual scientific evidence. Rather than just accept the evidence as-is, you try to rationalize that the evidence could be deceptive in nature. That’s not following the evidence, that’s sticking to your beliefs in spite of the evidence.
Rumraket,
I really think this claim is unsupported. What we observe like the nested hierarchy does not follow an ancestral pattern so closely that it is the only possible conclusion. Genes are not directly following an ancestral pattern. If God is showing anything it is that common descent as in universal common descent is false.
That isn’t even a grammatical sentence, but its core seems to be a claim that separate creation of species is incompatible with common descent of species. While true, it’s a tautology, i.e. a syllogism in which the premise is identical to the conclusion. And you have provided no evidence for the premise.
Can you explain what you mean by that? Is it still your misunderstanding of Sal’s flower?
What would count as evidence of special creation short of God appearing from the skies, talking in a booming voice and creating hierarchically arranged creatures before your eyes? If that is your requirement to believe, I respect that since I think being skeptical is a great quality.
However, short of that, is there anything in principle that would persuade you, like say the improbability of random mutational accidents creating coordinated post translational modifications (PTMs) not only in histone proteins (as shown by NIH ENCODE), but TopoIsomerase proteins (as I have shown above), and maybe the large majority of human proteins?
If you say the PTMs don’t persuade you, all you’re saying is that it won’t persuade you. Your criteria of what counts as evidence isn’t mine. You haven’t provided mechanism that explain the design of functionally coordinated systems like the PTMs in histones.
COMMON DESCENT IS NOT AN EXPLANATION FOR FUNCTIONAL DESIGNS.
John Harshman,
Not just Sal’s flower. Additional evidence that is confirming Sal’s flower which shows genes not following the tree and gene specific mutation rates (may not be mutation at all) that are not following the pattern.
Regarding the “flower” diagram, the term is “pan genome” diagram (I just learned the term myself).
Now regarding patterns of diversity and similarity across species of a protein.
If the Designer made all proteins the same across species the exact same way, we would have hard time elucidating functions, domains, and motifs. Right?
This addresses the question of the role of hierarchical patterns in biological discovery, btw.
If there are patterns of diversity, and deeply conserved regions, the deeply conserved identifies critical residues (like metal binding, i.e. the C2H2 zinc finger residues). Let us call these primary functions.
Now, if there are taxnomically restricted conserved regions, this can be used to identify secondary functions, like say species specific functions. Right?
Well well, I guess that tends to create hierarchical patterns in biology!
Look at the diagrams I provided. Look at the PTMs that tend appear in primate or mammal specific motifs. Heck if there is a lysine in mammals that don’t appear in fruit flies and yeast, there isn’t going to be a PTM at that location in fruit flies and yeast. Simple, and hierarchical.
I’d say that evidence of special creation would be species that didn’t connect in a nested hierarchy. Your problem is that we don’t see that evidence. You can ignore the evidence all you like, but don’t attribute your strawman skepticism to me.
Once again you confuse the evidence for common descent with the evidence for the causes of mutations. If (big if) you have evidence favoring divine intervention in the sequences of proteins and such, that is as compatible with guided evolution as with separate creation of species. Given that all the evidence is against separate creation of species, guided evolution would see to be your best hypothesis. Why do you reject it?
Still misunderstanding the issue, as has been your consistent practice from the beginning.
And nobody claims it is. That’s just you completely misunderstanding the issue again. Common descent explains nested hierarchy, i.e. the distribution of features. It doesn’t explain the origins of features.
Could you be more specific? Note that Sal’s flower does in fact support nested hierarchy, as I showed you quite a while ago, in that almost all the flower can be explained by a single change on the same tree. The tiny residue can be explained by two independent losses, which we expect to happen occasionally, but much less commonly than a single loss.
But beyond that, what would count as evidence? Apparently functions that can’t be the result of random mutation and natural selection don’t count as evidence for special creation in your book. Is that right?
No, I’m not sure we would. Conservation across species is not the only way to recognize function.
No, I don’t think it does. It addresses the question of the role of variation. You aren’t even looking at hierarchical pattern.
No, it can’t. Species specific functions would not, by definition, be conserved.
No, it doesn’t. Or wouldn’t. It would create species-specific differences; there’s no reason those differences would have a nested hierarchical structure. And there’s especially no reason for different loci to display the same hierarchical structure.
What do you mean by “mammals”? If there’s separate creation of species, there’s no such thing as “mammals” except as an arbitrary grouping of convenience. Why do we call whales mammals instead of fish and bats mammals instead of birds? Why do different loci group them the way they do? Your hypothesis explains nothing, while common descent is the very simple, obvious explanation for all that.
Of course it’s right. Why would you think so? I will point out that you have turned “special creation” into an ambiguous term, so it isn’t clear what you intend it to mean here. You might be referring to separate creation of species (standard creationism) or you might be referring to separate creation of changes in DNA sequence within individuals (guided evolution). The latter is not evidence for the former, and I wish you would stop conflating them.
Or if you think that non-random changes actually are evidence against common descent, please explain why.
Sure there would be because of patterns of similarity. Richard Owen’s definition of homology was based on common design, not common descent.
Unfortunately you don’t have any idea what you’re talking about, as usual. And you’ve shown no evidence of even being able to understand the points discussed on this matter. You are still apparently not even clear on what consilience of independent phylogenies even means, and you have been consistently wrong in your intuitions about what should follow from splitting lineages independently accumulating mutations.
What does that even mean? What does it mean to say, for example, that “a gene is directly following an ancestral pattern”?
Thank you for this completely unsupported declaration. Any idiot can do this. If God is showing anything it is that universal common descent is a fact.
Hey, it seems to work just as well in reverse.
So now we’re back to this: why should there be a nested hierarchical pattern of similarity? Why should that same pattern extend through morphology and genetics? (And why should it extend through stratigraphic and biogeographical occurrences too, for that matter?) You have no explanation for this consistency of a single nested hierarchy.
I see you’re back to your old pattern of ignoring most of what I say, including the central point, to concentrate on peripheral issues. Why is non-random change across species evidence for separate creation of species?
John Harshman,
John, you can explain any evidence that does not follow the pattern that you claim to prove the theory. This is evidenced by the evolutionists sweeping the Y chromosome difference with chimps under the carpet.
You only expect these things to happen because that is what you are observing. You do not have any evidence of why so many genes are lost or gained only that gene loss is possible so that is the explanation.
Well, along with other data I see evidence that the hierarchy you are claiming supports common descent doesn’t.
THAT’S FOR YOU TO FIGURE OUT FIRST.
You are trying to reject one testable explanation in favor of another explanation. But your other explanation needs to be a falsifiable scientific explanation that predicts particular patterns in the data in the same way common descent does. And preferably an explanation that is equally parsimonious and economic, while having at least equal and preferably better (otherwise why switch as you’d at best end up with a whitewash?) explanatory power.
But you don’t have such a model. You have a platitude, a phrase you never really define what the hell even means. You aren’t in possesion of a model just because you can type the words “common design” into a browser window. It becomes a model when it is turned into something that looks like a process, that has a mechanism, and from which particular outcomes should follow.
You might object that God is inscrutable, and the “mechanism” is when He Wishes For It Really Hard And Then It Just Happens And So Makes No Particular Predictions, but then that’s a problem for you, and we’re not going to throw away the scientific method because you want to have God inserted in science textbooks. Sorry.
Look, we can conclude this entire endeavor right here if you just admit that you’re not here promoting a testable scientific hypothesis, but rather your religious faith, your “assurance of things hoped for, the conviction of things not seen”. Then we can home. You will have advertised for you religion while not making a complete mockery of the scientific method.
I’m merely pointing out you are closing your mind to other lines of evidence of special creation such as improbability arguments.
I showed, if the Designer wants to created nested hierarchies just for kicks, it can be done. However, if there are systems that common descent cannot explain except by appeals to special creation of systems that define taxonomic groups, like say eukaryotic spliceosomes that define eukaryotes, then it’s rather unparsimonious to invoke common descent at all, one may as well invoke special creation of species.
So why did you avoid providing a simple “yes” or “no” to the question if other lines of evidence might or might not persuade you of special creation?
Is it because if you said, “yes”, then I’d ask why don’t you look more at improbability arguments. If you say, “no”, then it simply indicates you’ve closed your mind to other possible lines of evidence.
No. Why the hell would it?
How would the nucleotide sequence of, say, DNA polymerase III in humans and mice being completely identical somehow make it harder to figure out what DNA polymerase III does in mice?
Call me strange but I think if things are identical in different systems, that makes it EASIER to figure out what they do. After all, isn’t it reasonable to conclude that if they have identical constructions, they perform identical jobs?
Look at what your desire to advertise for a creationistic religious viewpoint have forced you to argue. How completely illogical and frankly ridiculous your suggestion here is.
Let’s just “test” this suggestion of yours on something designed with a little thought experiment. I give you two sets of electric kettles you don’t know how function in any detail, except in the superficial sense that you know electricity is converted to heat which boils the water. For each set, I give you the opportunity to take one of the kettles apart and analyze it for how it works and what each component does.
Now I give you the 2nd kettle from each set and ask you to do the same thing, take it a part, analyze it and determine what each component does.
Now in the first set, the two kettles are completely identical. How long does it take you to figure out what each component does in the kettle that is completely identical to the one you just took apart and figured out how works? I think it will take you exactly the amount of time it takes you to realize they’re actually identical.
For the 2nd set, the two kettles are of different designs and manufacture. They have commonalities to be sure, as both are obviously still kettles. But the electronics are wired differently, one possibly have more settings than just on-off, the locking and the mechanics of the lid-opening joint is different, and so on. How long does it take this time?
Look, I don’t think I have to say anything more here. What you’re suggesting is OBVIOUSLY LUDICROUS. Your prior commitments to the creationistic conclusion took you here, it forced you to make a COMPLETELY RIDICULOUS suggestion.
What are you talking about?
Once again you have confused the pattern, nested hierarchy, with the origin of the changes that make up the hierarchy. No matter how many times I bring that to your attention, you never learn. Why genes are lost or gained is not relevant to the fact that the pattern of gain and loss fits a nested hierarchy very well.
What is that evidence? I’ve asked you for that evidence, not a repeated claim that evidence exists.
Here is the next adjacent segment of Top2A and Top2B PTMs. Note that this segment has lots more non-phosphorylating PTMs. Is that by accident or design?
Click to ENLARGE
Conserved only has meaning when something is unconserved. So the answer is “yes” since the conserved regions often identify catalytic or metal binding residues.
Any bets where the metal binding residues of TopoIsomerase are conserved? Hmm, I’m in the process of posting TopoIsomerase alignments. Want to make a gentleman’s bet as to the level of conservation across species for metal binding residues? 🙂