
Dr. Winston Ewert put forward his module hypothesis, but I put forward an alternate module hypothesis at the domain and motif level of proteins. It is based actually on papers by evolutionists who have pointed out that the problem of “Promiscuous Domains” remains an unsolved problem in evolutionary biology.
When I put Promiscuous Domains on the table in the Common Design vs. Common Descent thread, the TSZ Darwinists ignored the problem and then declared victory. I viewed their non-response as evidence they didn’t understand the problem and/or preferred to ignore it.
Perhaps pictures are worth ten thousand words. From the NIH, that great source inspiration for the Intelligent Design community, we have the CDART database viewer.
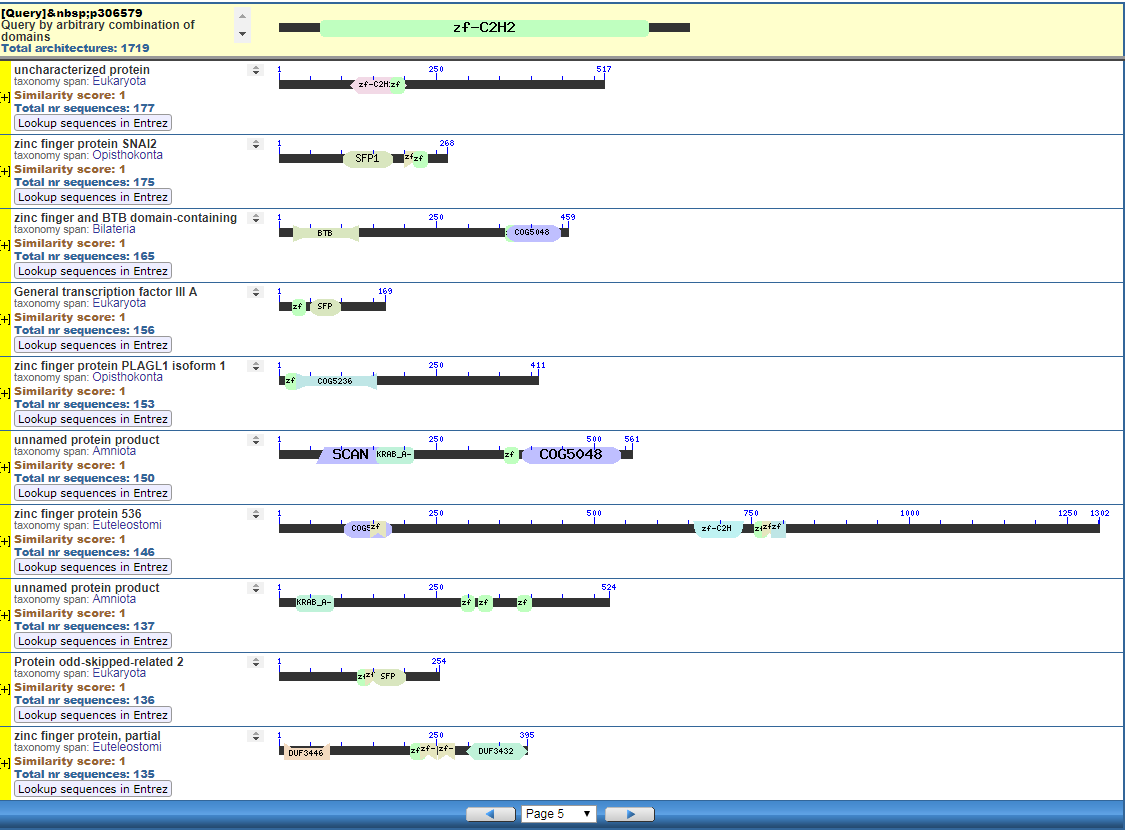
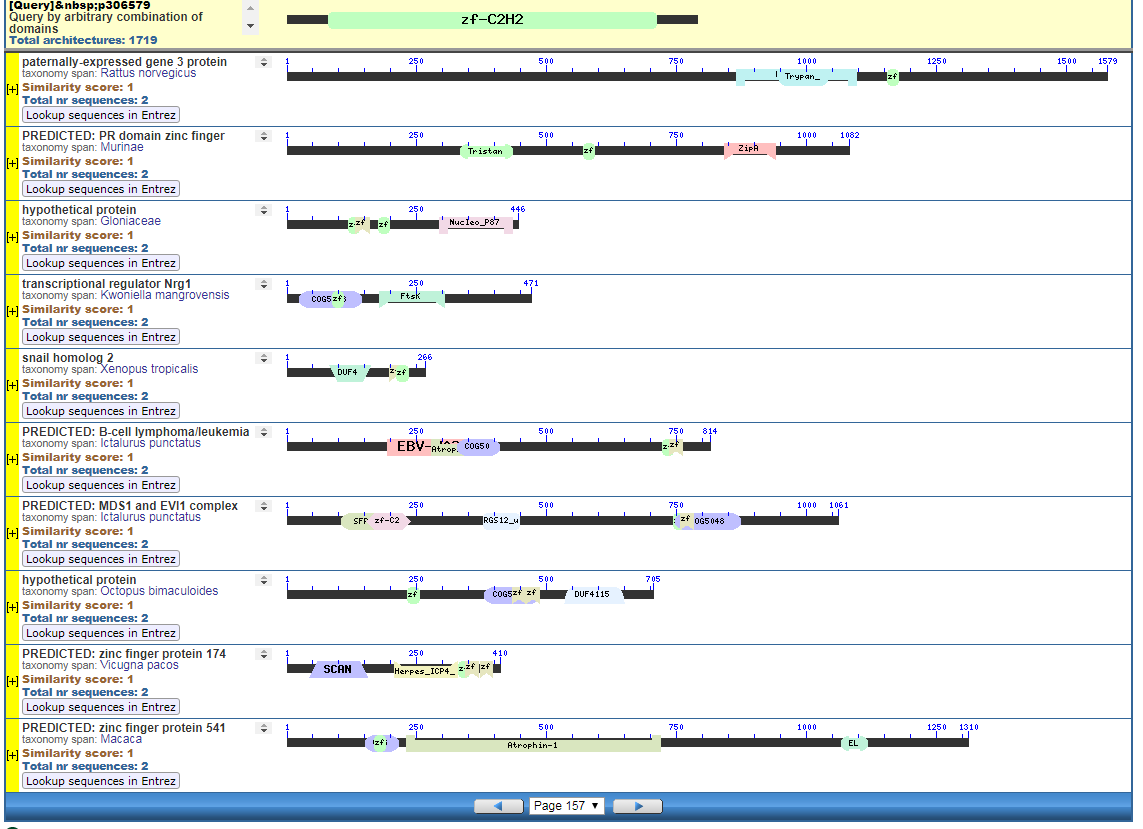
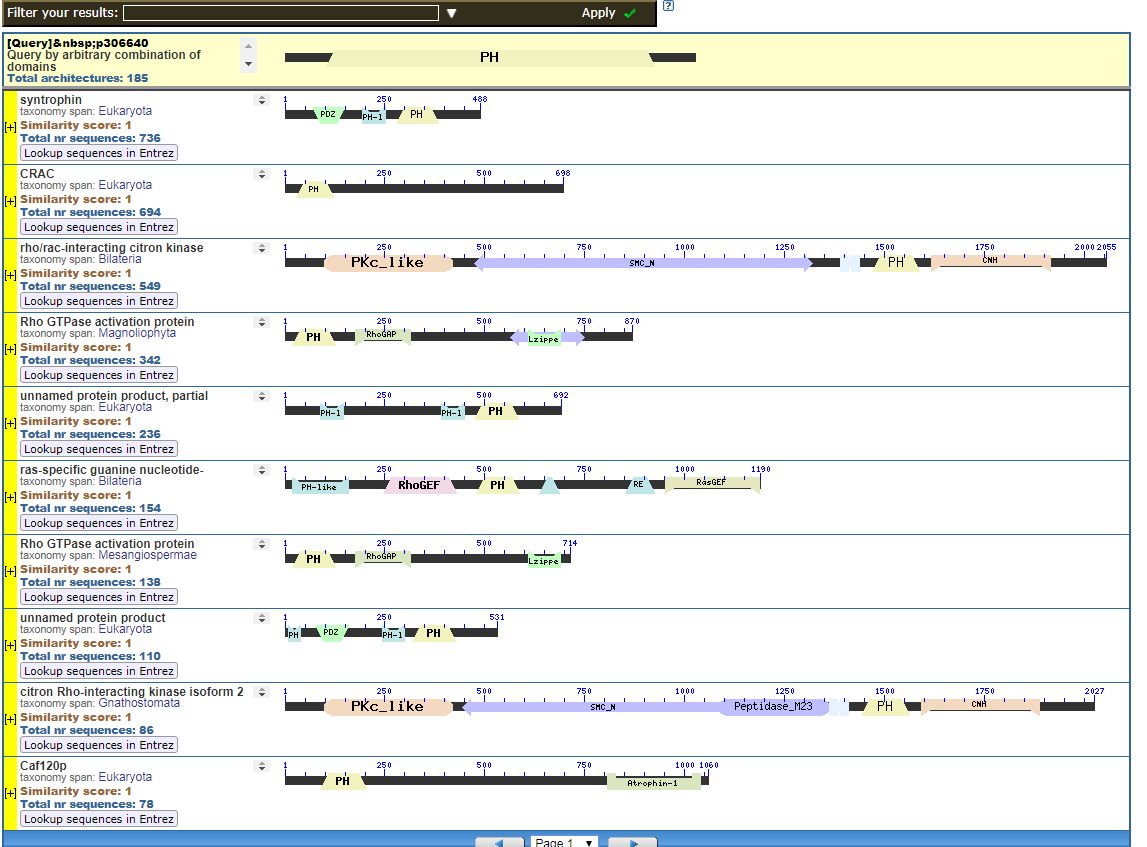
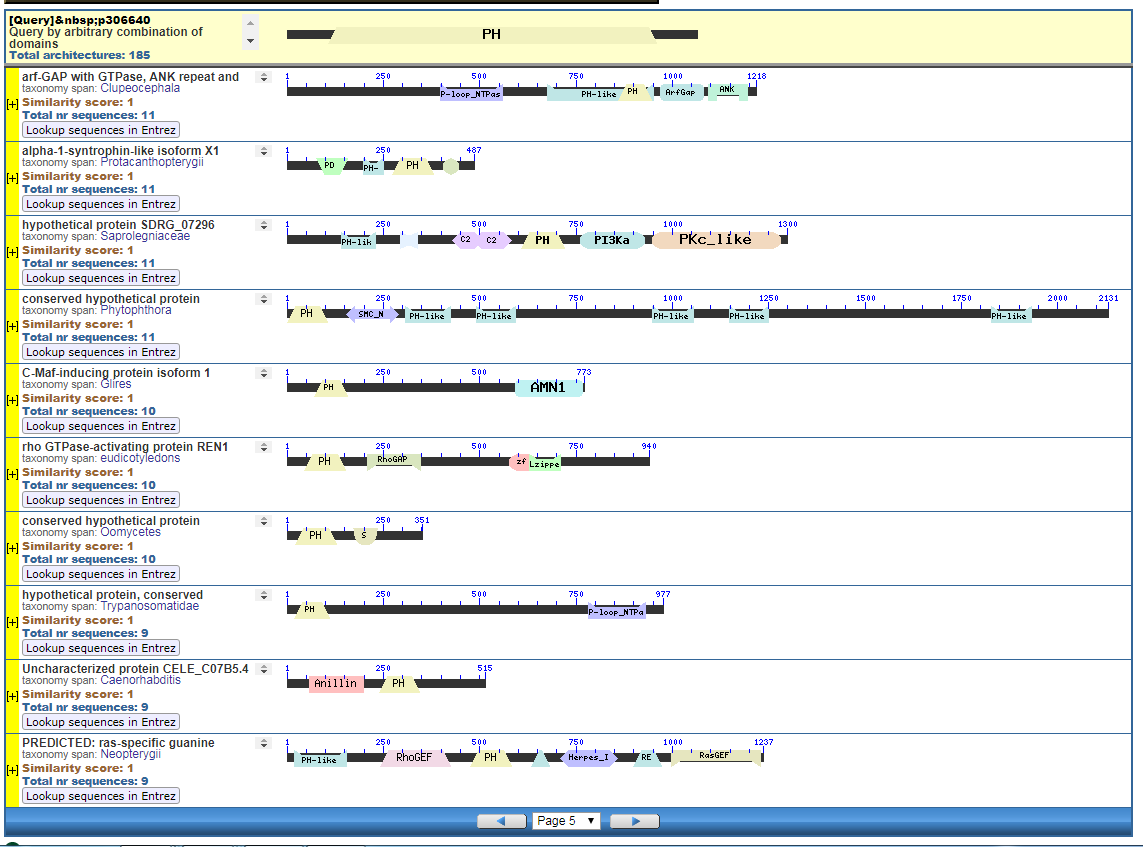
From the CDART viewer, I provide a few of the thousands of diagrams that show the promiscuity of protein domains. The diagrams below show the classical zinc finger ZF-C2H2 “ZF” domain and the Plextrin Homology “PH” domains. Note how the location of domains is “shuffled” to different locations in different proteins. It’s as if proteins are made by different lego-like parts in different order and position. My preliminary look into small 4-amino acid motifs that are the target of phosphorylating kinases suggests the the problem of promiscuity goes all the way down to small motif levels.
Such promiscuity is more consistent with common design than common descent.
Click to Enlarge Classical ZF-C2H2 Zinc Finger Page 5
Click to Enlarge Classical ZF-C2H2 Zinc Finger Page 157
Click to see all CDART Classical ZF-C2H2 Zinc Finger Architectures
Rumraket, are those PTMs functional or are they functionless chemical accidents?
Neither. They’re the result of mutations filtered by natural selection. Why won’t you understand that you’re engaging in a false dichotomy? In what language should we explain it so that you stop making such a basic mistake?
I don’t know what Rum thinks, but from here it looks like some might be functional, some might not be. Some might be mostly inconsequential. Why should this be an all-or-nothing situation? All you have is a multiple alignment. Don’t rush into hasty conclusions.
I don’t see the factual nature of evolution, nor even the possibility of design, as being necessarily tied to either of those options.
I think it could easily be the case that the majority of them are functional, and some few are nonfunctional but not disturbing enough to be visible to selection, and that they mostly evolved gradually over the entire evolutionary history of the proteins in question.
The fact of common descent is not contingent on a particular position with respect to the frequency with which putative phosphorylation sites (say) are functional.
I’m merely pointing out that the improbability of particular mutations has nothing at all to do with common descent of species.
You didn’t have to show that. It’s obvious that a designer (especially an omnipotent one) can do anything at all. But that to is irrelevant to science or to evidence of common descent, because it predicts nothing and forbids nothing. In other words, it’s vacuous.
Why? You have to present an argument that “special creation” of mutations is less likely than special creation of whole species. So far I see only your personal inclination to go that way. If you’re looking for a model that fits both the “improbability” data (if that really is data) and the nested hierarchy data, it seems that guided evolution is your only real choice. Why are you so resistant?
Because I already told you what lines of evidence might persuade me of special creation, and I already told you that your supposed evidence is irrelevant to the question. (And you are still using “special creation” ambiguously in a way that obscures the issue.)
Thanks.
Rumraket,
Woa!
Sure it does, because if features entail special creation, then there is no need to invoke common descent for those features. And if one starts invoking special creation of certain features, then why not all features? You’re only objection is, “I don’t see why a designer would make hierarchical patterns if he created things, therefore the hierarchical patterns were evolved, even though much of it could have been specially created.”
Seems to be more parsimonious to argue a tree and giraffe were independently created rather than trying to argue trees and giraffes had the same ancestor. I don’t believe in common descent anymore because it just seems silly to think a fish can give rise to a giraffe. Btw, such a transformation would entail lots of changes in PTMs.
I don’t think you appreciate the barriers that large scale changes in PTMs entail since this entails more than just mutating amino acids in a protein, it entails building “writers”, “readers” and “erasers” to process those PTMs. It’s not as trivial as supposing an amino acid simply mutates with no functional consequence.
Joe Felsenstein’s book on Phylogeny hardly, if at all, even addresses the problem of PTM evolution and transformation. PTM evolution entails COORDINATED simultaneous changes in proteins and protein interactome networks, not random uncoordinated mutation in individual proteins as most evolutionists assume.
Sal joins nonlin in doing “science by absurdity”.
Your god must be a super bore at dinner parties.
It’s always puzzled me why creationists like Sal posit “COORDINATED” changes when really nothing needs to be COORDINATED at all. It was created ex nihilo originally, why does the next version have to be connected to the last at all? Does god have a quota of what it can create whole and what it can push around via small changes in the genome?
Is that why we don’t see new species being created around us Sal, has your deity used up that quota already?
These absurd questions flow easily once you have follow Sal down that rabbit hole. That they are utterly unanswerable troubles him not al all I’m sure.
When we guess what his deity would do that’s absurd. When he does it, somehow that’s science.
I figured out what would count as evidence for me, but I was asking what would count as evidence for you?
If you saw God created before your eyes and speak from the skies would that count? That hypothetical question has relevance since, even Jerry Coyne said something like that would persuade him of a miracles. So, why is something like that believable as a miracle? Answer: God of the gaps. Gaps that aren’t a gap of knowledge but a real, or at least credible gap.
I’m pointing out the gap in the PTM and promiscuous domain evolution is not a gap of knowledge, but one of principle.
FWIW,
I’m making the above PTM diagrams for circulation in other venues. Since it seems most Darwinists here are totally interested in such amazing designs in biology, they are spending little time pondering the barriers to common descent it poses, unless one invokes miracles of special creation to rescue common descent, which is most ironic.
Anyway, I’d like to thank John Harshman for offering his views, though we disagree.
The OP was about promiscuous domains and motifs being better explained by common design. The PTMs in motifs spread across many proteins is a problem of promiscuous motifs. The above diagrams laid some of the ground work for search for promiscuous motifs. These motifs appear simultaneously in several proteins at once (or close to the same time). Such convergences are not consistent with common descent for the motifs, but such convergences in functionally coherent ways does not seem credible from random mutation.
There’s an impressive amount of things wrong with that statement as a reposponse to what I wrote.
First of all, conservation comes in degree, it is not completely conserved nor completely unconserved. Some things are over 90% similar between mammals and prokaryotes, other things have undetectable levels of similarity yet might still actually derive from a common ancestral sequence.
Second, the immediate implication is that you must think that the things which are at the lower, possibly undetected levels of conservation, were designed to be inscrutable in function by the designer?
Third, that you agree that conservation correlates, at least roughly, with the effect on fitness that mutations in the sequence has, and that therefore very unconserved regions are probably nonfunctional junk.
Fourth, like, what the fuck? Identical regions being identical and therefore can be used to figure out what they do in different organisms, does not hinge upon something else not being identical. Figuring out that the gearbox in car A is identical to the gearbox in car B and therefore (entirely reasonably) functions in the same way, is not in any way contingent on whether the wipers for the windshield are similar or wildly divergent.
Nothing you write makes sense even from whatever design perspective I’m trying to imagine you want to advocate for.
Are you asking because you think there is a level of conservation of those sites that would constitute an argument against common descent? My guess would be they are quite well conserved, not because evolution says so, but more because of the function of topoisomerases is well embedded in the processes of DNA replication and gene expression. I think it is generally the case that the greater the number of processes that depend on a particular function, the more conserved that particular function will be over generations. More dependencies -> slower change. Not necessarily zero change, just slower.
See? You’re conflating creation of species and creation of mutations under the common term “special creation”, which makes everything you say about it ambiguous. Common descent doesn’t explain the origin of any feature whatsoever, not just the ones you think can’t be natural. That isn’t what common descent is supposed to explain, so its failure to explain those features is not relevant to common descent. Common descent explains what it’s intended to explain, and what if true comprises evidence for it, e.g. nested hierarchy.
The objection to “the designer just happened to do it that way” isn’t that I can’t think of a reason why he would; it’s that invoking the whim of a designer can’t in principle explain anything, because it has no expectations for what we would or would not see. “It just happened that way” is an equivalent non-explanation.
We expect to see a nested hierarchy from common descent. We have no expectation of nested hierarchy from any other cause. Thus nested hierarchy is evidence of common descent. “Why not all features?” is not an argument.
“Seems to be” and “it just seems silly” are not arguments. Would you care to present an actual argument against common descent? I would be happy to consider it.
Are you claiming that changes in PTMs and the origins of “writers”, “readers”, and “erasers” all happen at the same points in the nested hierarchy? If so, what evidence do you have for that? All you have so far are mutations in amino acids at particular sites. Anyway, what does the complexity of a change have to do with evidence against common descent?
As it shouldn’t, since it has nothing to do with inferring phylogeny.
You have presented no evidence that this is true, and even if it were, so what?
I don’t know why you would thank me, since you have learned nothing from our discussion. You have never responded in any meaningful way to anything I have said. Cargo cult science it began, and exactly the same cargo cult science it continues to be.
What’s going on here is that Sal is deeply, deeply troubled by the nested hierarchy. He knows common descent is the only reasonable explanation and he’s struggling badly with that thought. I have no doubt in my mind that he, unlike Bill or Nonlin, is capable of understanding John’s argument that the common descent conclusion is independent of the mechanisms of variation and none of his “arguments” are relevant to the question of common descent. It wouldn’t be a huge problem if he wasn’t a YEC, but alas, he is.
No he doesn’t.
What would count as evidence that mutation would have to be coordinated simultaneously?
Actually common descent doesn’t explain taxnomically restricted features like spliceosomes in eukaryotes that have no credible ancestor in the fossil record nor even in principle. Nevertheless, such special creations of such systems will create nested hierarchies where common descent will fail as a mechanism to create such systems.
John Harshman seems fine with rescuing common descent with acts of special creation however. I think such a “fix” is unparsimonious.
can you name any process at all that produces categories that can’t be arranged in a nested hierarchy??
I certainly can’t think of anything
Everything from modes of transportation to musical instruments to geometric shapes to political parties can be arranged like that.
It seems to me that such an organization is the result of how we humans categorize more than something in the categories them selves.
peace
I don’t know, but I do know what would count as evidence that it was coordinated simultaneously: does it all occur on the same branch of a phylogenetic tree? Of course you would need pretty dense taxonomic sampling for that to be good evidence.
As I’ve said countless times and you never manage to notice, common descent doesn’t explain any features. It explains only the distribution of those features among species. Spliceosomes are no different in that respect from anything else. Special creations of systems will not create nested hierarchies unless whoever creates them makes a particular point of creating a hierarchy for no necessary purpose. Common descent is not a mechanism to create systems. How is it possible after all this time for you to retain such a fundamental confusion?
Once again, “I think” is not an argument. You have to explain why it is. As long as separate creation of species fails to explain the nested hierarchy — and it does — it can’t be parsimonious.
No. Anything can be arranged in a nested hierarchy just by declaring it so.
That’s because you have no understanding of the difference between arbitrary nested hierarchies and nested hierarchies that result from hierarchical structure of the data. Some sorts of data actually do have such structure, and it can be objectively demonstrated that they do. That’s true for phylogenetic data, but it isn’t true for modes of transportation, musical instruments, etc. Would you agree that if what I say here is true, then what you said is wrong?
You certainly need to explain what you mean by objective demonstration.
Any criteria that you wish to use will be the result of subjective choosing.
I can choose to group and categorize animals by size,color or by number of vertebra the choice is mine there is nothing objective about it.
I would agree that if you could in some way demonstrate that one persons choice of data for organization is objective and another persons choice is subjective then you will have accomplished something philosophically astounding.
I’m all ears
peace
To save us both some trouble here is a quote that purports to explain the difference between subjective and objective nested hierarchy
quote:
The difference between classifying cars and classifying languages lies in the fact that, with cars, certain characters (for example, color or manufacturer) must be considered more important than other characters in order for the classification to work. Which types of car characters are more important depends upon the personal preference of the individual who is performing the classification. In other words, certain types of characters must be weighted subjectively in order to classify cars in nested hierarchies; cars do not fall into natural, unique, objective nested hierarchies.
end quote:
from here
http://www.talkorigins.org/faqs/comdesc/section1.html
what you need to do is demonstrate that all observable features of organisms are given equal weight in constructing your hierarchy
good luck with that
peace
Nor even in principle? LOL.
Btw, there are no big protein complexes found in fossils. Ancestral states of the spliceosome are inferred by comparative genetics. Which points to group II self-splicing introns. But I guess you can just declare your own opinion as a matter of fact here: In your opinion you don’t find group II self-splicing introns to be a “credible” ancestor to the spliceosome.
Well in my opinion they are. And in my opinion my opinion counts for more than your opinion, so where do we go from here?
for instance I could group grey foxes together with grey squirrels and grey koala bears by color and climbing habit.
Would those “characters” be weighted equally with the ones you choose?
peace
Why would the spliceosomal complex require special creation? Why could it not have evolved from group II self-splicing introns through, for example, a neutral evolutionary ratchet ala the constructive neutral evolution suggested by Arlin Stoltzfus, Michael Lynch, and others?
Also, common descent explains the pattern in the distributions of spliceosomes(eukaryotes). As in, they exist in all eukaryotes because they were present (however they emerged) in eukaryotic lineages before the last eukaryotic common ancestor, and all subsequent eukaryotic lineages are derived therefrom and therefore contain spliceosomes. Common descent does not purported to explain HOW the spliceosome evolved. That would be the job of a theory of the evolution of cellular and molecular complexity such as this one How a Neutral Evolutionary Ratchet Can Build Cellular Complexity.
So rather than an occasional act of special creation (not that I agre that is required of course) that is subsequently inherited and undergoes slight subsequent divergence, every single thing must be an act of special creation, and that’s more parsimonious? LOL.
The next adjacent segment on Top2A and Top2B is below. The pattern does not look random and is clearly different than other segments in the distribution of PTMs.
Click to ENLARGE
I would need a model of special creation first before I could determine what evidence for that model looks like. You need to give a model of special creation, as in some sort of mechanism that entails particular patterns, before it can be determined what evidence for the model is.
I can’t just decide to consider some thing as evidence for special creation if I have no idea what would follow if special creation is true. I’m not the one claiming to know what a special creation model would entail, other than to say I think a special creation model that invokes deception in the sense that the creator is deliberately going out of it’s way to create the pattern that a common descent model predicts, is not a good model, as that pattern is already predicted by common descent so putting the special creator in as an additional factor is at the very least completely unnecessary.
Yeah if I saw God create something before my very eyes I would then believe God created the thing I saw God create before my eyes. Assuming I knew what God looked like. If stuff just appears out of thin air, but noone’s around doing anything, how am I supposed to conclude it’s God of a particular abrahamic brand doing the creating?
I still wouldn’t believe God created the diversity of life, unless I saw God actually do that (again, assuming I know what God looks like, which I don’t). God magicking some particular organism, or whatever other entity, into existence before my eyes would not convince me everything else was created by God too. That just wouldn’t follow.
Being persuaded that miracles can and do happen, would not suffice to convince me that everything CLAIMED to have happened by miracles, actually did. All claims must be assessed on a case-by-case basis.
God creating a fly does not mean God created all bears and horses. It just doesn’t follow. God walking on water does not mean God made you win the lottery. It just doesn’t follow. God turning water into wine doesn’t mean God personally intervened to get you off drugs. It just doesn’t follow. And so on and so forth.
God creating a bear and a horse does not mean God once took a fleshy form and called himself Jesus Christ. Every single one of those claims would require it’s own set of evidence and would have to be assessed on a case-by-case basis.
A horse zapping into existence before my eyes from a big hand coming out of the clouds does not somehow automatically entail literalist Biblical Christian theism of the young Earth creationist brand.
That makes zero logical sense.
So a gap of knowledge, the textbook fallacy. lol
You’re not “pointing” this out, you’re blindly asserting it is a gap. More importantly, you’re faulting a theory for not explaining something it isn’t even purporting or required to explain. Common descent does not make any statements about the frequency with which motifs or domains are supposed to evolve. It is merely supposed to explain why there is consilience of independent phylogenies, and why there is significant levels of hierarchical structure detectable in the data.
The explanations for particular features and entities, like spliceosomes, domains, motifs etc. are found in the theories of molecular evolution, the physical and chemical causes of various types of mutations, and population genetics.
You keep regurgitating this line that “the pattern does not look random”.
What does that mean? What is the implication of the pattern looking non-random and why?
For example, the concentration of phosphorylated serine residues localized to one region, that doesn’t look like result of random mutation.
What if God in the person of Jesus Christ told you that he created a bear and a horse and everything else?
This is important.
Most folks I know don’t reason from the creation of the bear to Jesus the Son of God.
They reason from the convincing and authoritative testimony of Jesus the Son of God to the creation of the bear
peace
How would you know it was the person of Jesus Christ? Because he said so? Because he looks like the guy in the Renaissance paintings? How? Why doesn’t God just implant the right thoughts in our heads directly?
How do I know you are the person of Alan Fox ??
something like that
peace
If you had the thoughts implanted then you would have no reason to commune with God and you would miss the entire reason for your existence?
peace
You don’t.
https://en.wikipedia.org/wiki/List_of_people_claimed_to_be_Jesus
You really can’t know anything at all given your worldview, can you?
peace
I don’t have any reason to commune with God as it is.
OMagain,
claiming to be Alan Fox or OMagain is not a guarantee that you are Alan Fox or OMagain
That sort of skepticism does not keep your family from being able to know when it’s you who tells them something
peace
Sure you do, You just don’t realize it yet.
peace
As you just once again demonstrated you need it so that you can know anything at all.
peace
This is rule-breaking, FMM. Please don’t assume what others think or believe. And we’re derailing Sal’s thread.
fifthmonarchyman,
Ditto.
I’m not assuming anything about what you think or believe. I’m assuming something about what we all need.
I do agree we are derailing the thread but you started it.
peace
I would be interested in knowing why you picked those two characters. Is it because you know I would consider the resulting classification bogus? And if so, would you agree that it’s bogus? I think you would, and that you think that foxes really ought to with foxes and squirrels with squirrels rather than declaring a category of “gray things”. And if so, aren’t you already supposing that classifications of this sort are not arbitrary and are in some sense “real”?
Now, I wouldn’t reject color as a character, but I would reject climbing habit, simply because a character has to be something that’s inherited. Color clearly is, but climbing habit is too vague a thing to be assured that it ought to be counted. I wouldn’t count the number of letters in the name either. Still, it wouldn’t do too much damage to include bogus characters as long as there were lots of others, and the characters were not chosen in order to favor a particular a priori notion of the story they ought to tell. Now, in practice, DNA sequence data are good because they are clearly inherited and because nobody chooses them based on their fit to a previously agreed on outcome.
Data can be shown to be objectively hierarchical if they haven’t been chosen for fit and if different subsets of the data (themselves not chosen for fit) give the same result. All the attempted hierarchies of transportation modes, etc., that I have seen are produced by picking one feature at a time, dividing on that basis, and further subdividing the divisions based on one feature at a time within that division. Pick different features, or even a different order of application, and you get a different classification. That’s by no means objective.
Alan Fox,
See folks from radically different worldviews can agree on things
peace
What does “look like” mean here, operationally? And is it on purpose that you talk here about mutation, full stop, and not selection too?
It’s because I like grey foxes and think the fact that they climb trees needs to be more well known.
Grey foxes and grey squirrels have much more in common than just color. They share similar habitat habit and diet and both are harassed by larger red members of their “genus”.
So there you have it, You admit you choose the character you choose because you think it facilitates the conclusion you wish to draw.
I’m tempted to rest my case right there
The problem arises when grouping based on DNA sequence do not match those based on some other character for example phenotype.
You just admitted that you choose for “fit”. in this case assumed heritable verses assumed not heritable So your point is moot.
Before we get too far in the weeds. I’m not denying the nested hierarchy and I’m not denying common decent.
What I’m denying is the “objective” business and the claim that special creation would not exhibit a nested hierarchy.
These are the things you need to prove
peace