
Evolution of KRAB Zinc Finger Proteins vs. the Law of Large Numbers
There are patterns in biology that violate the law of large numbers, and thus suggests Intelligent Design or at the very least statistical miracles. The pattern involves KRAB-ZnF proteins that have multiple zinc finger domains side by side that are inexact copies of each other and would require a scenario of co-evolution of their DNA binding partners with every additional zinc-finger insertion — a scenario indistinguishable from a miracle.
The role of a zinc finger can be something like a clamp or a lock. Each zinc finger in a KRAB-ZnF protein is fine-tuned to connect with a DNA much like a lock (the zinc finger) can receive a key (like DNA). Here is a depiction of a KRAB-ZnF protein with 4 zinc fingers as part of a large chromatin modifying complex. The four zinc fingers are marked “ZN” and connect to DNA:
The evolutionary explanation of side-by-side repeated patterns of zinc fingers such as in KRAB-ZnF proteins shows a severe lack of critical thinking by evolutionary biologists who pretend “phylogenetic methods” are adequate explanations of mechanical feasibility of common descent.
To illustrate the problem, consider the KRAB-ZnF protein known as ZNF136. For reference, this is the amino acid sequence of ZNF136:
https://www.uniprot.org/uniprot/P52737.fasta
This is an amino acid fragment contained within the ZNF136 protein
TGEKLYDCKECGKTFFSLKRIRRHIITH
This short sequence is called a zinc finger domain which in 3D looks like this:
https://sciencescienceeverywhere.files.wordpress.com/2015/08/figure-11.jpg
{kind=link}
Wiki gives a description of the function of zinc fingers in proteins that have them:
http://wikipedia.org/wiki/Zinc_finger
A zinc finger domain requires two “C” amino acids and two “H” amino acids placed in the right positions. It requires a few other things too…
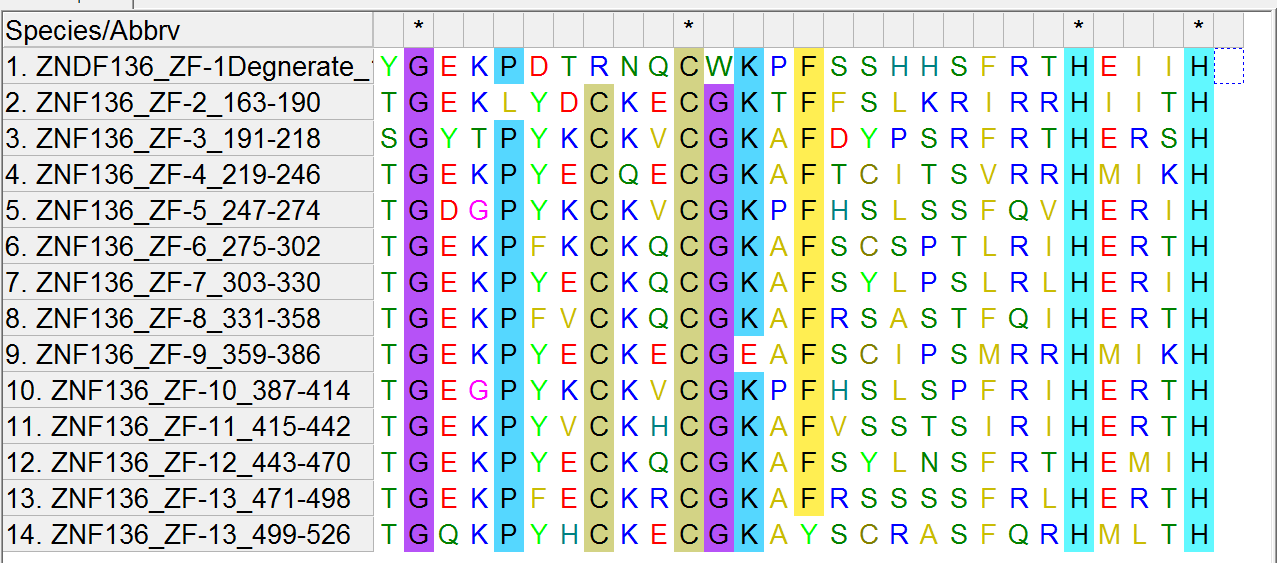
There are 13 zinc fingers in the ZNF136 protein and these are their sequences:
TGEKLYDCKECGKTFFSLKRIRRHIITH
SGYTPYKCKVCGKAFDYPSRFRTHERSH
TGEKPYECQECGKAFTCITSVRRHMIKH
TGDGPYKCKVCGKPFHSLSSFQVHERIH
TGEKPFKCKQCGKAFSCSPTLRIHERTH
TGEKPYECKQCGKAFSYLPSLRLHERIH
TGEKPFVCKQCGKAFRSASTFQIHERTH
TGEKPYECKECGEAFSCIPSMRRHMIKH
TGEGPYKCKVCGKPFHSLSPFRIHERTH
TGEKPYVCKHCGKAFVSSTSIRIHERTH
TGEKPYECKQCGKAFSYLNSFRTHEMIH
TGEKPFECKRCGKAFRSSSSFRLHERTH
TGQKPYHCKECGKAYSCRASFQRHMLTH
For completeness, there is a degenerate zinc finger in ZNF136 with the sequence “YGEKPDTRNQCWKPFSSHHSFRTHEIIH”
Why are the Zinc Fingers so different in sequence (except for the conserved amino acids)? To target a section of DNA, the zinc finger must be tuned to target it. Think of the zinc finger like a lock and DNA as a key that fits into the lock. In fact, for both the study of biology and medical applications, humans have a desire to make their own zinc fingers — like lock smiths. To bind a large segments of DNA, side-by-side zinc-fingers have to be tuned to their respective side-by-side DNA partners such as illustrated here:
By the way, there is a website that helps researchers construct the right amino acid sequence to make a zinc finger for a particular DNA target:
https://www.scripps.edu/barbas/zfdesign/zfdesignhome.php
Now, to visualize the critical/conserved amino acids, see the protein sequence here with highlights on “C” and “H” amino acids.
http://theskepticalzone.com/wp/wp-content/uploads/2019/04/znf136_zfC2H21.png
{kind=link}
Note one of the lines is not exactly like the other lines in that it is missing a “C”. This is the degenerate zinc finger mentioned above. So there is 1 degenerate zinc finger and 13 functional ones.
From this diagram it is apparent that the regular appearance of “C” and “H” is a violation of the law of large numbers, hence this pattern is not due to random point mutation alone. To “solve” this problem, Darwinist explain the pattern through segment duplication followed by some point mutation and natural selection, but this is not mechanically feasible either!
To understand why, let the reader first ponder the alignment I made of the Zinc Fingers in the ZNF136 protein using MEGA 6.0/MUSCLE software:
http://theskepticalzone.com/wp/wp-content/uploads/2019/04/znf136_zfC2H2_muscle1.png
{kind=link}
Then let the reader, consider the distance matrix generated by MEGA 6.0 which measures the number of nucleotide and percent differences between the zinc fingers.
http://www.creationevolutionuniversity.org/public_blogs/reddit/znf136_distance_matrix.xls
(All of the above results are reproducible, so I leave it to interested parties wanting to confirm the results to do so.)
For the duplication to work, at a bare minimum the right 84 nucleotide segment must be chosen, and then perfectly positioned for insertion so as not to break a pre-existing zinc finger. But supposing the duplication succeeds, why are the zinc finger’s conserved features involving “C” and “H” and other amino acids preserved and not eventually erased by point mutation given we obviously see the zinc fingers are different from each other. To preserve the “C” and “H” and other necessary amino acids in a zinc finger, the new zinc finger needs to be under selection. But in that case one is simply concocting a “just so” story for those newly minted zinc fingers without any respect for the difficulty of such a “just so” story being probable — and it is not probable!
To understand the problem of such a “just so” story, recall zinc fingers bind to DNA regions. Btw, this includes DNA regions such as ERVs! And ERVs are indicated to participate in the Stem Cell Pluripotency regulatory network:
https://www.nature.com/articles/nature13804
Further, the KRAB-ZnF protein is part of an incredibly complex machine that does chromatin modification (as shown above) by often attaching to ERV targets. But this would require that ERVs (or whatever DNA target) needs to co-evolve with the KRAB-ZnFs that attach to them!
Do these researchers even consider the fact such evolution would have to be instantaneous otherwise it would degrade function and not let the genome have the opportunity to adapt to the new accidental copy of the zinc finger because the duplication would immediately be selected against!
The above KRAB-ZnF complex is like a read/write head acting on Chromatin. Chromatin itself is an amazing mind-boggling design akin to computer ROM and RAM in one.
Again, the difficulty of evolution via random insertion/duplication mutations followed by point mutations is that such events would disrupt the binding of an already operational set of zinc fingers. For example, suppose we have an array of 10 zinc fingers side-by-side that collectively bind to a target DNA. Suppose one zinc-finger is duplicated and the number of zinc fingers is increased from 10 to 11. Oh well, the binding ability is broken or at best compromised, much like adding a single letter to a pre-existing password!
In sum, there is a violation of the law of large numbers in KRAB-ZnF proteins which is not explained by random mutation, nor random segment duplication followed by some point mutation and fixed by natural selection. Some other mechanism for the emergence of such proteins is indicated and would likely be indistinguishable from a miracle. Given the importance of such zinc finger proteins in the control of ERVs which are important in the stem cell pluripotency regulatory circuits, the origin of KRAB-ZnFs is even more miraculous.
Dang right, because it makes biology look evolved.
Allan Miller,
I agree, we at are at best in the first inning of a nine inning game.
Corneel:
Sal:
Why would that be bad news? (I know how most creationists would answer that question, but I’m curious to hear your answer(s).)
stcordova,
When you are ready Perry Marshall would be very interested in this work. One of his thesis is the world was designed for discovery.
The ground work has already passed peer-review several times starting in the 1970’s with Andy Wong. Wong became co-author with lead author Kirk Durson in 2012:
https://www.ncbi.nlm.nih.gov/pmc/articles/PMC3524763/
The project sat on the shelf for 7 years thereafter, and now in 2019 it’s getting revived because there was interest in the pharmaceutical quarters in Kirk’s improved version of Wong’s work. A team (which includes me) is now working on it, and the team is growing. My presentation is a bit of recruitment pitch for growing the team.
So, instead of seeing the diagram below through the lens of phylogenetics and common descent, it can be seen through the lens of a functional hierarchy. The argument is whether the nested hierarchy diagram below should be interpreted in terms of PHYLOGENY or in terms of FUNCTION. I argue it is better interpreted by function since the deviations cannot be random as a matter of principle:
Kirk took 1000 variants of ubiquitin sequences across species. He ran it through the algorithm described in his peer-reviewed paper with Wong. Below is the diagram you can find in his paper. The net result is the diagram highlighted chemical and structural features which correspond to what has been confirmed by x-ray crystallography of the proteins.
Many times it is difficult to get good x-ray crystallography of a protein, and it would be good if a functional approximation in terms of protein folding and chemical reactivity could be inferred from cross-species comparisons for other proteins that have yet to be characterized by crystallography or which cannot be characterized by crystallography.
Also, many functional proteins don’t really have much of a fold! Or they may get changeable folds based on post-translational modifications.
Kirk’s tool suggests the patterns of diversity and similarity are not the result of common descent and random mutation, but are patterns optimized for scientific discovery — but that metaphysical aspect is of little importance to the phrama/biotech/medical industry if the tool actually delivers in enabling structural and chemical predictions of proteins. But the metaphysical aspect is icing on the cake.
It would be bad news for creationists if we evolved since it would falsify sufficient (but not necessary) reason to believe in a Creator.
It wouldn’t necessarily be bad news for ID people like Mike Behe who accept common descent.
I used to be an evolutionist because of the nested hierarchy AND the progression of complexity through archetypal transitionals.
But superficial appearances can be deceiving like this pencil apparently bending by Snell’s law:
Maybe I am just stupid, but I don’t see how the consilience of independent phylogenies helps Kirk Durston discover stuff about the structure of ubiquitin. Could you explain?
Not seeing that. I understand that it would require some adjustment on behalf of creationists to find that humans evolved from more primitive ancestors, but how does that falsify sufficient reason to believe in a Creator?
Sorry for being a bit slow on the uptake.
This is where, finally, you will meet reality face first.
If YEC was true the creationists would be billionaires. They’d know where to dig for oil, right?
If your “team” get’s somewhere then we’ll be reading about you in Forbes. Or not.
The shame of it is that even commercial failure will not convince you. There will be some excuse as to why the fault lies elsewhere.
Yes, indeed. And the truth will eventually out, all the faster in a commercial situation.
First it would be helpful to consider what creationists (not necessarily anyone else) consider sufficient reason to believe in a Creator. There are creationists who, like me, are skeptical of the Bible being from God rather than fabrications of men….
Evidence life is young is sufficient reason for creationists to believe in a Creator. John Sanford’s work on Genetic Entropy is one example. Another evidence life is young is the graph below showing invariance in racemic chemical composition of amino acids, which indicates the fossils are the same age. I can explain if you want…
http://www.creation-science-prophecy.com/amino/racemization-rate.gif
But factual evidence contrary to creation is evidence there is no need of an Intelligent Designer therefore no need of a Miraculous Creator, therefore no need of God — at least as far as biology goes. Maybe in the Deist or cosmological sense there might still be a Creator, but if life evolved naturally, biology is not evidence of a Miraculous Creator.
Michael Behe is an example of someone sort of in between a Creationist and an Evolutionist. He’s one of the few centrists on issues that are usually polarized.
I’d say he’s more in the camp that common descent is true, but that it requires some form of unspecified design. His commitment to common descent is probably rather nominal, however. He mentioned in passing in Darwin’s Black Box that there could be front loading, akin to developmental ontogeny, that made common descent possible with common design. But he never developed that theory much. Mike Gene attempted to do so….
But, as far as me personally, and for other creationists like me, evidence in favor of common descent would make a lot of us agnostics or atheists or deists.
So, yes, you were spot on, the nested hierarchy is deeply bothersome to me. It’s the best argument for common descent and for evolution naturally bringing about the diversity and complexity found in biology. It’s a more powerful argument than natural selection.
Is it your position that there is zero evidence in favour of common descent then? As logically it seems it must be so.
Thanks for your answer.
Being a non-believer myself, it is somewhat hard for me to appraise, but I find it hard to believe that evidence of divine design in biology is the only thing that prevents you and other creationists from leaving the faith. Do you really suppose that you would be incapable of finding a way to reconcile common descent with your belief in God, when so many other christians do?
The nested hierarchy is not an argument, it’s very strong evidence for common descent and you know it
No it doesn’t. You’re just giving it that spin without considering it carefully. The idea of correlated positions came about in structural biology in the prior century. It was based on the expectation that inter-dependent residues, like those in contact, would tend to co-evolve, which is why the method is often called “evolutionary trace analysis.” These analyses have been performed since the early 90s (if not before), though the sequence information wasn’t enough for it to be as successful as it can be today.
The main group doing this kind of thing was Chris Sander’s. They published several articles actually solving 3D structures somewhat recently using evolutionary coupling (example here). The need for hundreds of sequences and for development of improved statistical methods, to get 3D structures, would be evidence that they’re far from being “optimized for discovery.” The same would be true about the noise in the data.
The co-evolution idea applied to RNA is the basis for some programs for predicting different kinds of RNAs, like tRNAs and rRNAs (with the infernal software). The results are heavily dependent on proper statistics. The statistical “boundaries” are diffuse. False and true positives mix up, suggesting that we miss a lot of real targets, and showing, again, that the divergences are evolutionary and far from “optimized for discovery.”
Anyway, good luck with the project.
Co-evolve? Like non-existent tansmembrane proteins co-evolving with membranes? Like random copy-and-paste-o-morphy mutations to co-evolve a topisomerase homodimer, forgetting that if it cut and failed to anneal, or cut and failed to do a DNA unwind, it would make a wreck of the genome.
The paper makes assertions without systematic consideration of the problems.
Feed yourself your own advice. My irony meter just blew hearing that come from you.
But as you refuse to enter that realm, of duelling papers, you stand mute.
Your criticism might well be on point but so what if nobody will ever hear it?
Strawman created and burnt. Check.
Well, I used to be a Christian evolutionist at one time. There was persistent belief in miracles and answered prayers in my family. We weren’t THAT strict about religion, but mom while living in a 3rd world country suffering hyper emesis through pregnancies was told to get an abortion. She and dad decided to carry the babies to term each time, but the 2nd time looked like it would be the end of mom’s life because she was so malnourished. Mom and dad prayed a lot, and my sisters were born, healthy and grew to be nurses. There were other incidents such as after Mom was in a car accident and the docs said she’d never walk again. The next door neighbor, a pastor, came and prayed for her, and there was movement in her leg that same day and she was able to walk again.
Sooo, even if I were convinced of common descent it’s possible to believe, but my sufficient but NOT NECESSARY condition for believing in God would be gone.
In other words, it is not necessary to reject common descent to believe in a creator, but if one rejects common descent based on one’s evaluation of the facts, one could believe in creation.
Alternatively, people like ID proponent Mike Gene, and perhaps indirectly Mike Behe, would regard origin of life a miracle requiring God, and then after the abiogenesis event, common descent happened via front loading.
Atheist biologist Jack Trevors, co-authored a paper as to why he thinks abiogenesis is outside of science and can’t be reduced to ordinary chemistry and physics. I concur with that. It’s sort of a “creationism” followed by common descent.
Common descent seems reasonable with like say close transitions like between man and other primates. But the gap between eukaryote and prokaryote, unicellular and animal life … those seem pretty unbridgeable as a matter of principle to me.
Something else there’s literally no evidence for despite decades of wishing hard.
So, for you the interventions happened in “real time”? At some point the universe was one way then it was different at some later point? The universe was created originally broken and has been “fixed” over and over again? Is that it?
Said this:
More here on Jack and his gang: https://sandwalk.blogspot.com/2008/04/guelph-creationists.html
I wasn’t quoting any paper. I was explaining that to you.
You did not read the paper(s) Salvador, otherwise you’d know that they had to confront the problems arising precisely from the imperfection of coevolving positions. Had those being “optimized for discovery,” the authors would not have had such challenges.
Well, what else can I expect? You miss explanations based in much more straightforward knowledge, no wonder you would not understand scientific articles.
It’s truly sad reading these things from you Salvador, it shows that you missed the whole explanation. My comment flew way above your head, as always.
stcordova,
That’s an awful lot of middle ground scooted over. You would be happy with the common descent of arthropods and birds, then? No? Spiders and flies? Weeeelll … Spiders and mites, then? Mites and ticks?
The problem is – for those not shackled by religion – that the assumed boundary does not manifest itself as the phylogenetic net is cast ever more widely. Relationship inevitably diminishes, but does not suddenly vanish at any remove. There is no firm basis for accepting phylogeny at one taxonomic level but rejecting the same methodology at another.
Actually Entropy, you’re right, I’m pretty much not reading your drivel and giving it much thought. Thanks any way for your links to the papers.
Oh, apologies to the authors. You were conveying your drivel.
Can you elaborate a little more. That was kind of cryptic.
Mutating zinc-fingers randomly is bad juju:
https://academic.oup.com/nar/article/45/3/1130/2574173
Oh well, Entropy and DNA_Jock proven wrong again by data. Any idea what proportion of random mutation of a zinc finger array within a transcription factor will help functionality? What are the odds of increasing function by getting the zinc finger to decrease binding to a functional location and instead bind more frequently to some random location on the genome or maybe not much of any place at all? 🙂
Did Entropy pump his speculations against the constraints of mutational probability, drift and selection within a population structure. NOPE! He just made a bald assertion. And when challenged on the details, he punts and says I don’t understand.
The problem is an acknowledge problem in evolutionary biology known as the waiting time problem.
Perhaps a 1-minute illustration of the Richard Dawkins wrestling with the waiting time problem might be helpful to Entropy:
A jocular, onomatopaeic reference to your tendency to Gish-gallop.
Thanks for sharing that. Yes, that was the kind of thing I was thinking of; Solace and hope and participation in a close community. Whether species share common ancestry or not seems such a trivial thing to lose one’s faith over.
That’s a surprising statement. The transition from an ancestral primate to modern humans is the one thing that most other creationists seem to be hung up on. Could you envisage yourself accepting common descent of humans and apes at some point, while rejecting the major transitions you just mentioned?
But species NOT sharing a common ancestor is definitely not a trivial thing for some to lose their (atheist) faith over.
Clearly that is the motivation behind the fanatic Darwin devotion.
That doesn’t seem right.
The real issue here is independent creation. For creationists, the problem with common descent is that it is incompatible with divine independent creation of modern taxa (that is my understanding at least).
Speaking as an atheist, organisms not universally sharing common descent wouldn’t really bother me. So life started multiple times? Oh cool! However, I guess that if species turned out to be independently created in their current form, that would be somewhat of a poser. I don’t think the facts support the latter scenario BTW (no surprise, I take it).
Many a night have been spent by all the world’s atheists, lying awake in puddles of sweat, tortured by the prospect that slime mold isn’t related to toe fungus.
What reason could there even be to live?
Rumraket,
+1. I’m crapping myself, frankly. Nothing would get me through that church door faster than an exception to the (so far) consistent pointers to LUCA being a genuine coalescent for all life.
Exactly, which is why you end up ridiculing yourself by pretending to answer things you didn’t read. You didn’t read the very article that you linked either, since they cite articles by Lichtarge about the evolutionary trace method.
Well, since you don’t think that you deserve respect, you reap what you sow.
Oooh yes, the projection is strong with this one.
Also safe to say that stcordova has not actually read the article cited (Barrera et al, 2016), nor pondered on the existence of over 50,000 human polymorphisms that affect DNA binding of Transcription factors (60 heterozygous and 20 homozygous per genome: that’s a LOT of bad juju!).
Oh boy.
I firmly believe in the second coming of LUCA, who will bring about a new tree of life to this depraved world. Remember only observants of the Word of Darwin will adapt and survive, while the rest will be unable to outcompete the 10 plagues of LUCA’s descendants and will surely face extinction
You’ve all been warned
The bold is in the original, and it’s very important to remember that. Some students in my genetics courses make the very same mistake that Salvador is doing. So, here a reconstruction of the common conversation:
Student: I found that all mutations in this gene are deleterious.
Entropy: Interesting, where did you find this information?
Student: In the Mendelian Disease Database.
Entropy: OK, what’s the name of the database again?
Student: The Mendelian Disease Database.
Entropy: What’s that word after “Mendelian”?
Student: Disease.
Entropy: So, what kinds of mutations are bound to be in the database?
Student: Deleterious mutations.
Entropy: What kinds of mutations would very rarely make it into the database?
Student: Ah! Oh! Of course, neutral and beneficial mutations would not make it into the database!
[pause to allow the other students to understand and consider what just happened]
Entropy: We don’t need to stop there. Do you think that neutral mutations are visible in the phenotype?
Student: Of course not! They’re neutral, they would be phenotypically invisible by definition.
Entropy: What about semineutral mutations?
Student: Not that much of a phenotypic effect, or not something to worry about.
Entropy: So, how likely it is that scientists in general, and the Mendelian Disease Database in particular will collect those mutations?
Student: Not very likely.
Of course, my students are willing to learn, and cannot pass the course without reading for comprehension. I warn them from the very beginning that shallow reading won’t do. Salvador? He’s not likely to read this and understand it.
It seems like they proved that you lack a bit of logic. “A bad mutations database has bad mutations! Therefore all mutations are bad!” It’s like saying, “A baseball stamp collection has men who are baseball players! Therefore all men are baseball players!”
Actually, yes. I do that all the time. I also consider the frequencies of gene duplications and recombinations, as well as the biochemical and biophysical phenomena that help understand both the constraints and the flexibilities in mutations, drift and selection.
You have never challenged me on any details. In order to do that you would have to first understand the explanations. For example, you still don’t understand ectopic recombination, otherwise you’d understand why it would naturally produce tandem multiplications with “perfect” boundaries. It’s obvious from the very process, and you keep missing it. You keep asking about the “perfect boundaries,” then you ask how can it happen “13 times!!!!!!!!” (exclamation points and all). Therefore, you don’t understand. No way around it.
Which is an imaginary problem that could only come from someone who has no idea about biochemistry, as myself and DNA_Jock have explained. DNA_Jock produced a whole OP explaining this, and you keep missing all of the points.
Will you make it this far? Will you try and understand? or will you continue ridiculing yourself? I think the answer is ridiculing yourself. Feel free to continue proving my point.
Yep.
Entropy,
Thanks again for the papers by Sanders. I’ll mention it to Kirk when I see him.
Ok, so explain to the readers the probability a gene with a zinc finger will make a zinc finger repeat. Like in a genome of 3.3 giga bases and a population of 7 billion people, how many times do you expect a pre-existing zinc finger to be duplicated, just once.
Alternatively you can express it in terms of new zinc finger repeats per generation.
Then answer how many zinc finger repeats might be damaged by a tandem repeat. Do you have an estimate.
Answering that question is more of an explanation than the hand waves you’ve given.
Anyone convinced that Entropy is right should be waiting for Entropy’s answer.
Everybody who’s convinced that I’m right, has already understood the answer Salvador. Otherwise they would not be convinced.
The right question is whether you’ll read it for comprehension if I answered this time, rather than ignore it, then pretend to answer while showing that incomprehensible cluelessness that characterizes you.
Since you missed this one, and then skipped this part in the comment you pretended to answer:
I doubt both your will and your abilities to understand. (Maybe you’ve got it and don’t want to admit that you’re logic is a tad problematic.)
So, if I expanded on the zinc finger explanations, would you try and read for comprehension for a change, or would you continue ridiculing yourself? Again, I’d bet on the latter.
I asked Entropy this question which he didn’t answer:
If he doesn’t have numbers, he can just say so. He doesn’t have to make the issue about me or my supposed lack of understanding.
Are we talking 1 per human per generation on average? 0.1, 0.001, …..
One measly number would suffice.
Let me appraise readers of a tactic if one wishes to avoid discussing inconvenient FACTS and concepts.
One thing to do is attack the participant. Cast aspersions to his supposed lack of understanding, etc. Talk about his motivations, his character, his education, his qualifications, whatever.
But, sometimes all that is being asked is to provide numbers for the probability that a certain class of events actually happened.
That’s the playbook if one’s goal is to discredit someone and thus take focus off of reasonable SKEPTICAL questions about claims made by evolutionary biologists.
At issue is the probability of evolving and FIXING functional proteins into a population. The thread isn’t about me, my qualifications, my understanding, my education, my background, my motivations, my character, etc.
stcordova,
There’s a lot wrong with your questions Salvador, but, again, we cannot go there unless you will read and try to understand the answers. I asked if you were going to read with the intent of understanding. Your avoidance to answer the question speaks volumes: You’d rather not promise anything because you know you lack the will, and maybe you also know that you lack the ability.
Here’s a question for Sal: is the probability of an initial duplication the same as that for an additional copy? Justify your answer.
For a solitary zinc finger in a protein, what is the probability that zinc finger will duplicate. You can state the parameters under which you can expect your figure to be valid.
You’re the one claiming ectopic duplication will do the trick.
No. Especially if they are identical.
But if they are identical tandem copies, then one can suggests some odds.
From said article:
Don’t know why you guys are celebrating. You think that’s evidence of good juju or at best neutral juju? That’s not exactly favorable to your objections to the problems raised.
I described my opponents playbook here:
Entropy plays it out:
You’re so predictable. Hilarious.
So how about some numbers. You name the parameters for your numbers. You could be direct and say you don’t have figures. But will you just repeat this sort of play?
My point is related to misidentification of homologous sequence during recombination. If you have two nearby regions with high similarity, this can cause copy number increase, with excellent boundary preservation. There is a mechanistic reason for this. It’s also one of the reasons transposons can cause problems, i.e. the probability of such misalignment is pretty high, given the basic condition.
Now you’ll leap on this and say ‘but it’s damaging’, and of course often it is. But you need it to be always damaging. It is widely accepted that most mutations are not beneficial.