
Evolution of KRAB Zinc Finger Proteins vs. the Law of Large Numbers
There are patterns in biology that violate the law of large numbers, and thus suggests Intelligent Design or at the very least statistical miracles. The pattern involves KRAB-ZnF proteins that have multiple zinc finger domains side by side that are inexact copies of each other and would require a scenario of co-evolution of their DNA binding partners with every additional zinc-finger insertion — a scenario indistinguishable from a miracle.
The role of a zinc finger can be something like a clamp or a lock. Each zinc finger in a KRAB-ZnF protein is fine-tuned to connect with a DNA much like a lock (the zinc finger) can receive a key (like DNA). Here is a depiction of a KRAB-ZnF protein with 4 zinc fingers as part of a large chromatin modifying complex. The four zinc fingers are marked “ZN” and connect to DNA:
The evolutionary explanation of side-by-side repeated patterns of zinc fingers such as in KRAB-ZnF proteins shows a severe lack of critical thinking by evolutionary biologists who pretend “phylogenetic methods” are adequate explanations of mechanical feasibility of common descent.
To illustrate the problem, consider the KRAB-ZnF protein known as ZNF136. For reference, this is the amino acid sequence of ZNF136:
https://www.uniprot.org/uniprot/P52737.fasta
This is an amino acid fragment contained within the ZNF136 protein
TGEKLYDCKECGKTFFSLKRIRRHIITH
This short sequence is called a zinc finger domain which in 3D looks like this:
https://sciencescienceeverywhere.files.wordpress.com/2015/08/figure-11.jpg
{kind=link}
Wiki gives a description of the function of zinc fingers in proteins that have them:
http://wikipedia.org/wiki/Zinc_finger
A zinc finger domain requires two “C” amino acids and two “H” amino acids placed in the right positions. It requires a few other things too…
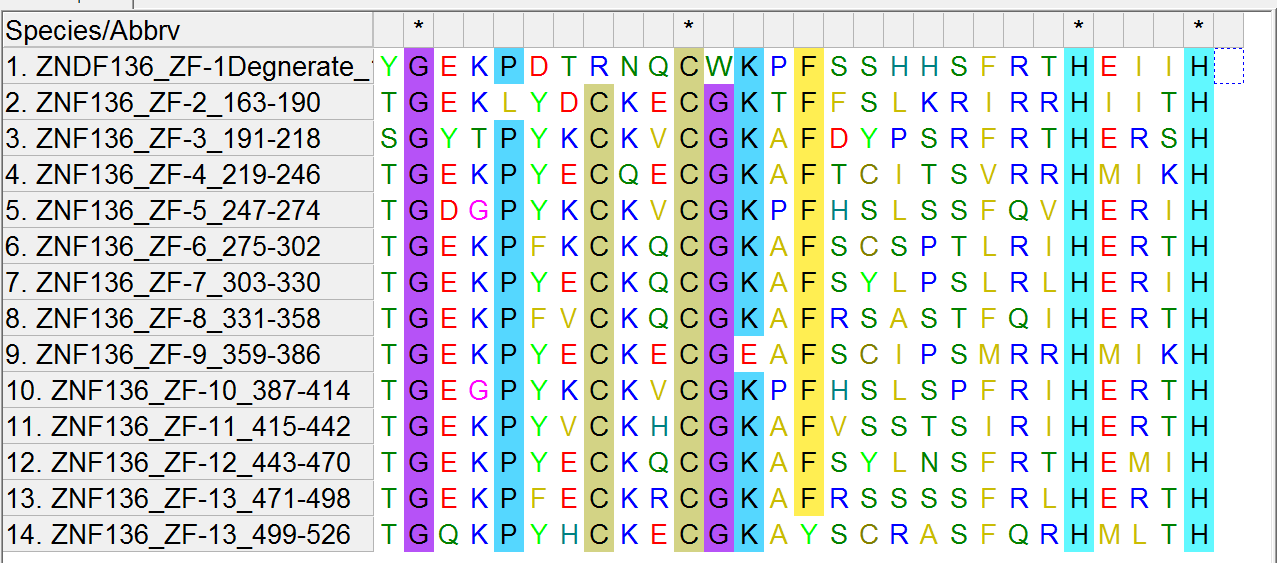
There are 13 zinc fingers in the ZNF136 protein and these are their sequences:
TGEKLYDCKECGKTFFSLKRIRRHIITH
SGYTPYKCKVCGKAFDYPSRFRTHERSH
TGEKPYECQECGKAFTCITSVRRHMIKH
TGDGPYKCKVCGKPFHSLSSFQVHERIH
TGEKPFKCKQCGKAFSCSPTLRIHERTH
TGEKPYECKQCGKAFSYLPSLRLHERIH
TGEKPFVCKQCGKAFRSASTFQIHERTH
TGEKPYECKECGEAFSCIPSMRRHMIKH
TGEGPYKCKVCGKPFHSLSPFRIHERTH
TGEKPYVCKHCGKAFVSSTSIRIHERTH
TGEKPYECKQCGKAFSYLNSFRTHEMIH
TGEKPFECKRCGKAFRSSSSFRLHERTH
TGQKPYHCKECGKAYSCRASFQRHMLTH
For completeness, there is a degenerate zinc finger in ZNF136 with the sequence “YGEKPDTRNQCWKPFSSHHSFRTHEIIH”
Why are the Zinc Fingers so different in sequence (except for the conserved amino acids)? To target a section of DNA, the zinc finger must be tuned to target it. Think of the zinc finger like a lock and DNA as a key that fits into the lock. In fact, for both the study of biology and medical applications, humans have a desire to make their own zinc fingers — like lock smiths. To bind a large segments of DNA, side-by-side zinc-fingers have to be tuned to their respective side-by-side DNA partners such as illustrated here:
By the way, there is a website that helps researchers construct the right amino acid sequence to make a zinc finger for a particular DNA target:
https://www.scripps.edu/barbas/zfdesign/zfdesignhome.php
Now, to visualize the critical/conserved amino acids, see the protein sequence here with highlights on “C” and “H” amino acids.
http://theskepticalzone.com/wp/wp-content/uploads/2019/04/znf136_zfC2H21.png
{kind=link}
Note one of the lines is not exactly like the other lines in that it is missing a “C”. This is the degenerate zinc finger mentioned above. So there is 1 degenerate zinc finger and 13 functional ones.
From this diagram it is apparent that the regular appearance of “C” and “H” is a violation of the law of large numbers, hence this pattern is not due to random point mutation alone. To “solve” this problem, Darwinist explain the pattern through segment duplication followed by some point mutation and natural selection, but this is not mechanically feasible either!
To understand why, let the reader first ponder the alignment I made of the Zinc Fingers in the ZNF136 protein using MEGA 6.0/MUSCLE software:
http://theskepticalzone.com/wp/wp-content/uploads/2019/04/znf136_zfC2H2_muscle1.png
{kind=link}
Then let the reader, consider the distance matrix generated by MEGA 6.0 which measures the number of nucleotide and percent differences between the zinc fingers.
http://www.creationevolutionuniversity.org/public_blogs/reddit/znf136_distance_matrix.xls
(All of the above results are reproducible, so I leave it to interested parties wanting to confirm the results to do so.)
For the duplication to work, at a bare minimum the right 84 nucleotide segment must be chosen, and then perfectly positioned for insertion so as not to break a pre-existing zinc finger. But supposing the duplication succeeds, why are the zinc finger’s conserved features involving “C” and “H” and other amino acids preserved and not eventually erased by point mutation given we obviously see the zinc fingers are different from each other. To preserve the “C” and “H” and other necessary amino acids in a zinc finger, the new zinc finger needs to be under selection. But in that case one is simply concocting a “just so” story for those newly minted zinc fingers without any respect for the difficulty of such a “just so” story being probable — and it is not probable!
To understand the problem of such a “just so” story, recall zinc fingers bind to DNA regions. Btw, this includes DNA regions such as ERVs! And ERVs are indicated to participate in the Stem Cell Pluripotency regulatory network:
https://www.nature.com/articles/nature13804
Further, the KRAB-ZnF protein is part of an incredibly complex machine that does chromatin modification (as shown above) by often attaching to ERV targets. But this would require that ERVs (or whatever DNA target) needs to co-evolve with the KRAB-ZnFs that attach to them!
Do these researchers even consider the fact such evolution would have to be instantaneous otherwise it would degrade function and not let the genome have the opportunity to adapt to the new accidental copy of the zinc finger because the duplication would immediately be selected against!
The above KRAB-ZnF complex is like a read/write head acting on Chromatin. Chromatin itself is an amazing mind-boggling design akin to computer ROM and RAM in one.
Again, the difficulty of evolution via random insertion/duplication mutations followed by point mutations is that such events would disrupt the binding of an already operational set of zinc fingers. For example, suppose we have an array of 10 zinc fingers side-by-side that collectively bind to a target DNA. Suppose one zinc-finger is duplicated and the number of zinc fingers is increased from 10 to 11. Oh well, the binding ability is broken or at best compromised, much like adding a single letter to a pre-existing password!
In sum, there is a violation of the law of large numbers in KRAB-ZnF proteins which is not explained by random mutation, nor random segment duplication followed by some point mutation and fixed by natural selection. Some other mechanism for the emergence of such proteins is indicated and would likely be indistinguishable from a miracle. Given the importance of such zinc finger proteins in the control of ERVs which are important in the stem cell pluripotency regulatory circuits, the origin of KRAB-ZnFs is even more miraculous.
Tally so far to my question:
Allan Miller: No.
Entropy: No.
I thank them for taking time to respond. I guess then all those hidden Markov Models of Theobald that Keiths swears buy have limits in proving common descent from a single ancestor from the very beginning of life, maybe from LUCA, but not from PUCA.
Nobody’s HMMs are supposed to “prove” that all proteins have a common ancestor, and life’s universal common ancestry doesn’t require that all proteins descended from a single one. If you thought that was the goal of some HMM, or that proteins had to have their origin in a single one for there to be universal common ancestry for extant life, then you’re very deeply confused. I’m not going to ask how you’ve got that weird idea because I doubt I’d get a coherent answer.
ETA: I’m not surprised by your confusion, and I’m not surprised that you really don’t care why there cannot be universal common ancestry for all proteins. But both facts are very telling.
So where did all the major protein families evolve from? Eh, like out of nowhere — ex nihilo.
This question doesn’t explain why you’d think that HMMs are supposed to “prove” that all proteins evolved from a single one. This question doesn’t explain why you’d think that universal common ancestry of all proteins is required for the universal common ancestry of all life either.
P.S. Of course not ex nihilo Salvador. You’re the creationist remember? Not from a single ancestral protein either. If you understood a tiny bit of molecular biology, or at least one percent of the quotes you’ve mined, you’d know why that’s just not possible.
Non-coding DNA, perhaps?
Even if all proteins really descended from a single ancestral protein, it would be near-impossible to detect this with the techniques available. Shortish stretches of genome are subject to far more signal degradation than longer ones – even though longer ones are made up of shorter ones. That’s the LLN at work. Additionally, pretty much every amino acid can be substituted, and it is probable that the early genetic code had few assignments, swamping the ur-protein signal with these later additions.
So even if it weren’t possible for common motifs to arise from random intergenic space (it is), the absence of that common descent signal is not evidence against it. Not that I think it is even there. What is remarkable, though, is the extent to which the patterns do accord with a hypothesis of organismal common descent, despite many aeons of signal degradation in the deepest branches.
I was asking because you never said whether you believe KRAB zinc finger proteins to be part of those “cellular processes that you cannot mess with to any degree”. If so, then they should be highly preserved, but that’s not true for all of them. If some of them belong to the collection genes that can be modified, degraded or broken, then Sal’s “bad juju” argument doesn’t generally apply.
Does that make sense to you?
Haha, do you have some first-hand experience?
Thank you for keeping the conversation alive.
Just 10 years at the ID-face! The classic opening gambit: “Speaking as an engineer, I …”. 😀
Sal, regarding your question of a single ancestral protein: What Allan said.
Salvador,
The root of the scop database classification is not evolutionary, but structural. The classification has both evolutionary and structural hierarchies because the authors wanted a complete scheme. While this would be obvious to anyone who knows a bit about structural biology, I don’t expect you to have knowledge about everything. However, since you want to make fun of evolution, you should be much more careful and avoid hasty conclusions. Maybe you should have asked someone with the appropriate knowledge to clarify for you if that root was supposed to be evolutionary before attempting to make fun of it.
Of course, you don’t have to trust me. Also, given your MO, you’d surely would start hunting for misquotes, or poorly understood quotes. So, I’d rather allow one of the main authors to explain to you, here, that the higher levels in the classification are structural and do not imply homology.
You’re welcome.
So for each of these it is your opinion that they are the direct, first generation products of design and that no explanation for their origin or any precursors can possibly be provided, even in principle?
I take it then that your “designer” can intervene whenever if feels like it and change our reality? Rather then, for example, it all being front loaded into the big bang?
Just noted that we have covered one of those before:
Mensur Dlakić and Arcady Mushegian (2011): Prp8, the pivotal protein of the spliceosomal catalytic center, evolved from a retroelement-encoded reverse transcriptase. RNA. 2011 May; 17(5): 799–808. PMID: 21441348
OMagain,
I think design is the best explanation for the origin of most proteins in general. We know they require complex sequences (DNA) and we know mind can generate complex sequences.
Corneel,
Did the paper include the design hypothesis as a possible alternative to the story it generated.
This like all other papers of its ilk assume protein evolution is true and then try and find similar sequence fragments in the past as evidence they evolved.
The problem is there is no explanation how from that historic fragment a new protein was formed.
When you find a historic protein to prpf8 that only takes a couple of mutations to form prpf8 then I will grant you a plausible path.
Most? So there are exceptions? Can you be more specific? Name a protein that evolved?
And were proteins designed before or after the big bang? E.G. did your designer design them at the start (front loading) or intervene in real time?
Perhaps I misunderstood. Weren’t those supposed to be examples of “very unique sequences”?
And yet:
You have no explanation other then “design” as to how a new protein can be formed. And “design” is not an explanation at all. And there is no possible pathway to such an explanation either.
For evidence, notice how you simply ignore my question as to when the designer intervened to create these proteins you claim can only be designed. Unless you decide what sort of designer you are looking for how can the search for design “explanations” begin at all?
OMagain,
Why are we even talking if you just want to make conversation stoping assertions?
Design is a process that a mind can make. It is a limited explanation however that does not mean it is not the best in certain circumstances.
Corneel,
We are talking about sequences whose origin is difficult to explain with proteins that exist in older species.
It’s only a conversation stopping assertion because it’s true.
In case you have not noticed, I’m asking you to demonstrate that it’s not true. For example which is more likely in your opinion and why: protiens were designed prior to the big band or they were designed in “real time” by interventions when they were needed?
The answer to that question goes a long way to deciding what your next step will be as an ID researcher.
It’s on par with “unicorns designed the first proteins”. It adds nothing to our understanding and it allows no further paths for investigation. As demonstrated by your inability to go beyond an assertion that “some things were designed”. You refuse to even take the next step and state what sort of designer you believe in.
Have you tried “their origin is that they were designed”?
Are you starting to see the problem yet? If Intelligent Design “wins” will the university textbooks simply be
Question: What is the origin of X?
Answer: It was designed?
Page after page of “it was designed”?
After all these years don’t you have anything else?
Why would anybody do something that philosophically incoherent? Scientists keep themselves strictly within the confines of science. fantasy cannot be included in their analyses.
Why shouldn’t they assume that protein evolution is true? There’s plenty of evidence for protein evolution after all. Scientists work with what they have. Give them actual evidence for magical beings in the sky and they’ll include them.
Let’s suppose that’s so. There’s still loads and loads of evidence for the evolution of plenty of proteins, and no reason to think that less studied proteins didn’t evolve. We don’t start imagining that some just-discovered-planet is moving around its star pushed by angels, rather than orbiting by gravitation just because the planet was just discovered.
The similarity is enough to suggest a path.
It’s amazing that you’d be this strict for natural phenomena, while being standard-free with fantasies as explanations. That doesn’t go well among scientists though. Among scientists you have to keep your standards coherent.
I don’t think he’d agree. I mean, it’s true, but nonetheless…
Design requires much more than a mind. It requires energy, tools, raw materials, processed materials, knowledge about such materials, bodies that respond to those minds and that have the dexterity for moving, assembling, etc. Design often requires teams with expertise in each part that will be assembled into a design. A mind itself requires complex biological phenomena just to exist, starting with energy to sustain the mind working, besides cellular, molecular and biochemical moieties and interactions.
So, before suggesting design as an explanation to something, you have to ensure that you can point, with confidence, to those designers and their requirements. Talking about a “mind” as if that encompassed everything that’s required for design to occur is ridiculous.
OMagain,
Of course he won’t agree. But one good day Bill might understand where he gets it wrong when talking to someone whose stance is scientific, rather than religious.
According to the paper, group II introns (the presumed precursors of our own spliceosomal introns) are the most likely origin of the reverse trancriptase in the core region of the ancestral Prp8. Does that qualify as a plausible path? Perhaps you feel that explanation is more cumbersome than “a mind made it”?
OMagain,
Ok. This is legitimate complaint but if we know the conclusion is tentative as all science is then it is fine. Design is the best explanation for the origin of life and unique species now. It may be replaced in the future with a more detailed explanation as our understanding of how.
There are many thing that design is not the best explanation for as more detailed explanations exist.
DNA and its functional sequences is very strong evidence of design as is the electron.
It’s no explanation, whatsoever!
Was that life created directly by what we would call an intervention? Was something created that was not there before that had no precursors? Or was it part of the design from the beginning, built into the big bang? Or something else entirely?
For someone who has a downer on ‘conversation-stopping assertions’, that’s a doozy. And wrong. Because of course, it’s the worst explanation. Especially when you leap from protein sequences to electrons.
Allan Miller,
We could talk about what a unique species is. Behe would define it at the family level and above.
Ok will retract the electrons for now but they seem pretty useful for something that is the product of a random accident 🙂
OMagain,
All great questions but above my pay grade 🙂
Alan Fox,
Repetition is good. I finally realize this is your opinion 🙂
Interesting. I’d think that if there’s no explanation, detailed or not, the answer is “I don’t know.” To Bill if there’s no detailed explanation the answer is “it’s designed.” So, until you know the details it was designed.
The best of this is that if you get explanations, it’s enough to claim that the details aren’t enough to fall back into “it was designed.”
I don’t know why I have this diarrhea. It was designed. Hey, you had Salmonella! Yeah, but the explanation is not detailed enough, therefore it was designed. I don’t know why that kid got cancer, it was designed too, etc.
I really don’t think he would.
In this case, it seems necessary, though Entropy has written the reply for me:
Interesting. I’d think that if there’s no explanation, detailed or not, the answer is “I don’t know.” To Bill if there’s no detailed explanation the answer is “it’s designed.” So, until you know the details it was designed.
The best of this is that if you get explanations, it’s enough to claim that the details aren’t enough to fall back into “it was designed.”
I don’t know why I have this diarrhea. It was designed. Hey, you had Salmonella! Yeah, but the explanation is not detailed enough, therefore it was designed. I don’t know why that kid got cancer, it was designed too, etc.
Thanks, Entropy!
Alan Fox,
Right. The cancer was not designed, it was selected for, obviously.
No problem.
Of course, because it’s obviously advantageous.
Ah, another dichotomy.
Actually I wasn’t confused, I asked the question to point out to Bill that even you guys don’t believe in universal common descent in places where it counts, like the origin of proteins.
I was just illustrating YOUR conflicted claims.
LOL!
The SCOP claim is the structural classification gives evolutionary relationships because the sequences CANNOT!!!!!!
Besides YOU, yes YOOOOOU, said all proteins didn’t descend from a common ancestor anyway. So my point was proven via several independent routes, not the least of which your own rejection of common descent of proteins. Thank you very much.
But the structural classifications fail too.
This is a listing of the major protein families according to SCOP.
The membrane proteins? Ah yes, an interesting class that has is also a challenge for common descent on many levels. More in my subsequent comment.
For the reader’s benefit here are diagrams of the locations of the cell membrane. There are also membrane bound organelles in Eukaryotes which I’ll describe later. But this is a diagram I will refer to in subsequent comments.
Unless common descent doesn’t require that proteins always descend from other proteins, in which case there’s no conflict.
Hey, nice to hear from you. Thanks for your comment.
Alan Fox,
Are you demonstrating to me that Entropy doesn’t yet understand the design argument? He has an excuse as he has not been posting very long.
Imagine if we humans didn’t have a mouth to take in food, water. Or a mouth and nose to take in and air — or just as bad, what if we didn’t have openings to get rid of waste! YIKES!
Well…there would be analogous problems for the first life if it didn’t have transmembrane proteins to allow the right stuff to go in the cell and the waste stuff to go out.
Roughly speaking the transmembrane proteins serve as gateways from the inside of the cell to the outside of the cell. They allow transport of the right substances and signals in and out of the cell. It’s important the right things go in and out!
Having no transmembrane proteins in the first cell is bad juju.
The membrane is made of lipids — a lipid bi-layer.
For reference:
https://en.wikipedia.org/wiki/Transmembrane_protein
But a picture is worth a thousand words. Note the different kinds of transmembrane proteins. It’s a stretch to suggest that all transmembrane proteins descended from a common ancestral transmembrane protein.
Of course, we could ask Entropy, Allan Miller, DNA_Jock, Rumraket, Keiths, etc. the following question:
So, the first cellular life needs transmembrane proteins to survive. They didn’t descend from pre-existing proteins, they sort of just popped up out of nowhere, or at the very least abiogenesis believers must claim unprovable, untestable, unrepeatable, unknowable, mysterious mechanisms to explain the emergence of trans membrane proteins in the first life — how is this much different than creationism, except that abiogenesis pretends no miracles are needed, but without justification.
Here is the picture. It’s blurry, if someone can find a better one, thank you in advance!
https://i.pinimg.com/originals/55/0c/7e/550c7eedaee7fcfbf4044ab75c6e6980.jpg
A challenge that evolutionary biology rises to. Remember, Sal, evolutionary biology and biologists don’t claim to have explanations for everything. There are models that sometimes don’t explain every detail. In those cases, biologists say “we don’t know” (see above). The way to supplant evolutionary biology as a model is to provide a better model. A model that explains the observed facts and makes testable predictions. “You can’t explain it therefore Design” won’t do that for you.
Salvador,
You’re taking things out of context (how surprising). Some structural analyses can help in determining common ancestries not evident otherwise. But those are tertiary-structure comparisons accompanied by biophysical, biochemical, and statistical analyses. However, not all structural classifications are supposed to reflect common ancestry. Some can be done merely for convenience, like the higher hierarchies in the SCOP classification scheme (as they said themselves).
Had you been careful, and followed the link I provided, you could have saved this spectacular, capitals-with-multiple-quotation-marks-and-LOLs, embarrassment (taken from here):
You could have read that yourself Salvador. I suggest that next time you read carefully before adding that many exclamation marks and “LOLs” to your comments.
Yes, MEEEEEEEEEEE. I said that not all proteins come from a common ancestor, which is compatible with what I told you about SCOP. SO what’s your problem then? Why won’t you be careful then? Why insist that SCOP fails on something they didn’t intend at all?
What point would that be? That not all proteins share common ancestry? Lots of scientists already know that, and, unlike you, lots of scientists understand why. We don’t need you to tell us.
I think what’s proven, yet again, is my point, since you keep demonstrating that reading for comprehension is not your forte.
Oh!? So that failure wasn’t enough? You want to keep adding to the evidence that you cannot read for comprehension? That you prefer to embarrass yourself further than accept a little tiny correction? Let’s see that then.
Really Salvador? You say protein families and give me a list of classes (according to SCOP)? Didn’t you read above? Here it goes again: “Classes based mainly on secondary structure content and organization.”
Seems like what fails is your ability to reading for comprehension.
As I said, and as the authors of SCOP said, these classes are based on some structural feature, not implying homology. So you’re telling me what I already knew, and what SCOP stated. Of course membrane proteins belong into different families, and of course not all of them have a single common ancestor. No need for you to explain this. We already know. Calm down.
Now, please. For your own sake. Stop and think.