
This is the third and last of my answers to Jonathan Wells’ 10 unanswerable questions for evolutionists.
Question 3: Homology.
Wells: Why do textbooks define homology as similarity due to common ancestry, then claim that it is evidence for common ancestry — a circular argument masquerading as scientific evidence?
This question stems from confusion on Wells’ part between how something is defined and how it is recognized, which are two quite different things. Homology is indeed defined as similarity due to common ancestry. But we don’t just label any similarity a homology and call it evidence for common ancestry. That would indeed be circular. What we really do is quite different. Similarity between the characteristics of two organisms is an observation. If the similarity is sufficiently detailed (“both are big” or “both are green” won’t do) we consider it a candidate for homology.
Homologies can be tested to some degree by predicting that the characters will be similar in ways we haven’t yet checked. For example, if we propose that similar-looking bones in two animals are homologous, we might predict that they would arise from similar precursors in the embryo, have similar spatial relationships to other bones in the organism, and have their development influenced by similar genes. And this is commonly the case.
But the main way of testing candidate homologies is by congruence with other proposed homologies. By congruence we mean that the two characters can plausibly belong to the same history. If the history of life looks like a tree, with species related by branching from common ancestors, then all true homologies should fit that tree; that is, each homology should arise once and only once on the tree. If a large number of functionally and genetically independent candidate homologies fit the same evolutionary tree, we can infer both that the candidates really are homologies and that the tree reflects a real evolutionary history.
And in fact that’s what we commonly find. Mammals, for example, are inferred to descend from a common ancestor because they all have hair, mammary glands, and other more obscure characteristics like seven neckbones and three earbones. All these characteristics go together: mammals have all of them and no other animals have any of them. Further, other characters support consistent groupings within mammals, and groupings within those groupings. Within most of life, groups are organized in a very special way called a nested hierarchy. In a nested hierarchy, every group is related to every other group in one of two ways: either one group entirely contained within the other (as in a below), or they share no members at all (as in b below). No two groups can partially overlap (as in c below).
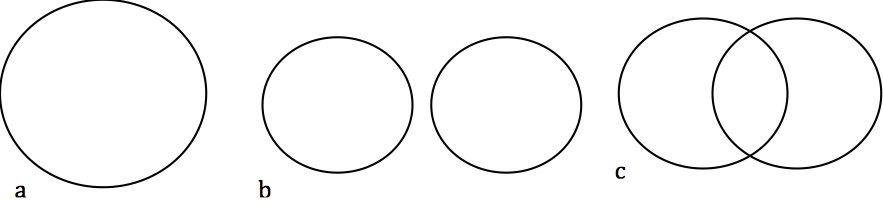
What we see if we try to organize species using candidate homologies is that groups organized according to different characters fit together like a and b, but not c, so we get a pattern like this:
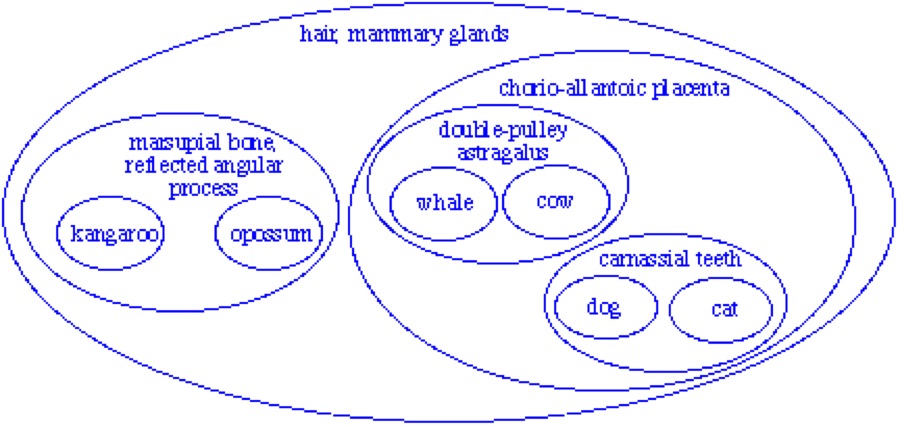
Why should these and many other characters all go together in this consistent way? Evolutionary biology explains these characters as homologies, all evolved on a single tree of descent, like this:
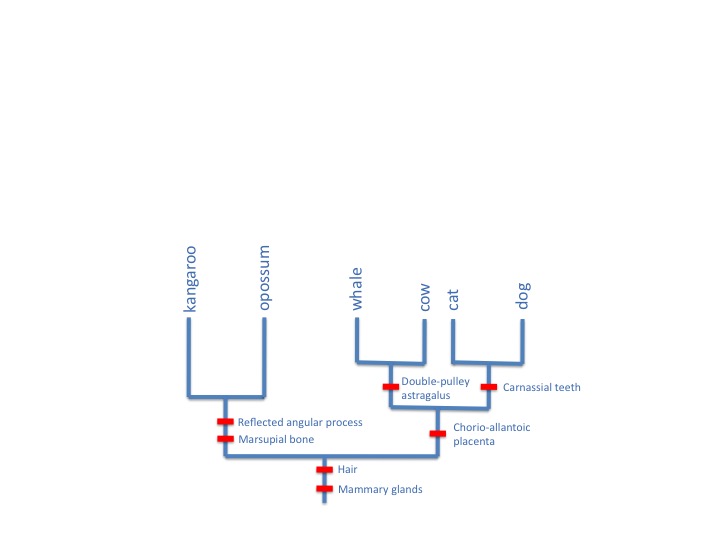
Wells gives no alternative explanation for such patterns, and indeed they are hard to explain in any other way than as reflections of an evolutionary history. Wells has it all wrong. Homology isn’t a circular argument, it’s a branching tree of evidence.
Of course homologies were recognized before they were routinely understood to be due to common descent. But only common descent actually explains such otherwise inexplicable similarities (design might do something like it in some cases, but hardly as extensively and in the patterns that we find in life).
Yes, unsurprisingly, homologies are defined by their cause, and someone like Wells just claims that it’s an assumption based on nothing but his ignorance. Wells is the kind of person that shows what a corrosive effect creationism has on an otherwise smart person. He went through college-level education in evolutionary science and largely failed to learn anything at all well about it. Denial is a strong barrier to learning, as also indicated by more local IDists/creationists.
Glen Davidson
The success of anatomical and embryological homology is so neatly confirmed by molecular phylogeny. Two independent approaches producing the same results!
Souls have a sort of “magnetism”, which unlike normal magnets have more then just two poles. They have multiple poles. Much like those personality tests you may have seen, but in many many more dimensions. And these poles interact with each other in more complex ways then mere repulsion and attraction. Like souls attract, and this can repel other ‘soul-groups’. This is partly why extinctions happen – souls are returned to an earlier stage of the materialisation process due to natural sorting in the pipeline, and when an animal is born and no suitable soul is available obviously it’s not going to survive to adulthood.
As these souls are allocated to the earthly plane they naturally sort themselves into groups. These groups are defined by the natural sorting that occurs when the souls come together. This is why mammals have characteristics not found in any other animal, it’s simply because in mammals the individual “poles” are so strong they push away any other animal with similar features back in the queue for instantiation. They exist, but we’ve just not been around long enough for them to come through yet, and they may not until existing mammals go extinct (as above).
The DNA you end up with is a by-product of the soul’s specific configuration. It’s the effect, not the cause. Observed evolution in the fossil record is expected with this theory as the poles are not static, they interact with each other in complex overlapping ways with multiple levels of complexity. This is also reflected in the resulting DNA on the earthly plane, as the DNA is a product, not the cause.
OMagain,
I’m expecting a strong endorsement from Charlie, but perhaps you could put in a word or two about etheric forces just to help him along.
Ah, I’m currently in my cardboard pyramid, I’d have to change over to the copper one for that!
Alan Fox,
Oh, like organisms need like DNA, don’cha know. Every last indel. And both will organise into a tree pattern ‘cos … ‘cos … ‘cos that was known before Darwin had even invented evolution.
In this passage you suggest that the main way of testing candidate homologies is by congruence, i.e. a co-existing set of homologies. This is why it seems to me that later in this passage there should be more “congruence(s)” instead of “homologies”.
For example, If the history of life looks like a tree, with species related by branching from common ancestors, then all true homologies should fit that tree; that is, each congruence should arise once and only once on the tree.
Right?
Just trying to help. This stuff seems worth it to make sense of.
No. Homology is the correct word. Congruence is being used here to refer to a match in pattern among multiple characters. A homology refers to a single character or evolutionary event. An evolutionary event happens once, at a particular spot on the tree, and the character is inherited by descendants of that spot. (Of course one event may resemble other events; if similar characters arise from different events, that isn’t homology, it’s homoplasy or convergence.)
John, you are right that the argument is not circular. It would be circular only if “common ancestry” were defined in terms of “homology” and it isn’t.
Let me give an analogous case.
I find “gill” defined on the web as the paired respiratory organ of fishes and some amphibians, by which oxygen is extracted from water flowing over surfaces within or attached to the walls of the pharynx.
But suppose I shorten it to Organ used by some animals to breathe under water.
Then I say that I’ve found a bunch of animals with gills and that is evidence that animals live under water.
That is perfectly acceptable so long as “animals that live underwater” is not defined as animals having gills. And it’s not.
Exactly the same thing is going on here. Wells is mistaken: as you correctly point out, there has been no arguing in a circle here.
“But the main way of testing candidate homologies is by congruence with other proposed homologies.”
Let’s say homology refers to what you say it refers to, but in context you seemed to be saying that to test whether it’s a homology you need congruence. From this, I assumed that to determine whether a character really is inherited (and thus can be called a homology, as opposed to chance resemblance), it’s actually the congruence that must be inherited along the line.
Where am I going wrong?
Hard to tell. Perhaps you are using the wrong definition of congruence. It’s a relationship between patterns. Relationships between patterns aren’t properties of lineages and can’t be inherited. It’s the homology that’s inherited. Now, homology is used in at least two senses, as a relationship between characters and as the character possessing homology (a homologous character) itself. I’ve tried to suggest the two usages by putting an article before the word or using the plural when I mean the latter.
Homology is similarity due to common ancestry. Congruence is a relationship between two or more candidate homologies, i.e. their fit to the same tree.
And by congruence you know they are homologies, and without congruence they are merely candidates, right?
As I said in the OP, there are other tests, but congruence is the main one. If you look at the third figure, which I wish had been bigger, “reflected angular process” is congruent with “marsupial bone”, since both are present in kangaroo and opossum but not in the other mammals. This is evidence that the reflected angular process in the kangaroo is homologous to the reflected angular process in the opossum and that the marsupial bone in the kangaroo is homologous to the marsupial bone in the opossum. Is that clearer?
So congruence of those characters (namely, “reflected angular process” and “marsupial bone”) in opossum and kangaroo tells that those characters are homologies and thus opossum and kangaroo have a common ancestor? And that ancestor had the same characters/congruence?
Erik,
Yes, except that you still seem to be using “congruence” incorrectly. A single ancestor can’t have congruence. Congruence describes the distribution of characters among multiple taxa.
And of course the conclusion of homology, like everything in science, is provisional. I wouldn’t want to rely on just two characters for any major degree of certainty.
It would be neatly agreeing if genetics followed body plans from a common design plan.
Its what a creationist would predict and desire.
We would not desire different dna for same loooking oparts on different creatures.
Eyeballs all get the same dna score based on a common design.
Its just a guess it shows common descent but no evidence except for the reasoning behind it.
its all comparativeolgy.
John harshman.
Welcome thread.
your very wrong on a point here. Wells is right.
Homology, a real biological fact, is said to be based on common descent and then they use likeness to prove common descent as the reason for homology.
It is circular reasoning if one is investigating WHY there is homology.
Common design easily is a option for homology and so it nullify’s homology is rightly defined as from common descent.
yoiur side is proving common descent by homology after it fIRST defines homology as from common descent.
We win this point.
All you can do is prove YOUR hypothesis for homology.
Homology belongs to mankind and all/any hypothesis. NO not just common descent. NO you can’t then prove common descent by homology BASED on common descent presumptions.
Evolutionism makes a great point for creationists.
Wells makes a good point. Give him credit or prove why he is wrong.
John Harshman.
Just a few points.
First you presume a tree exists and then you make your predictions for how things should fit.
Yet a non tree but common design concept would also predict these things. Including branching into species.
You bring up mammals. I say there is no such division in Gods mind or nature called mammals.
its just people, weirdly matching up traits. ALL of them are obscure traits.
there is no reason that mammary glands, hair, etc should connect creatures unless you know it already.
Its just good ideas based on a creators plan.
Why not do it that way?
A female rhino and a female mouse and our mothers having mammary glands is not evidence for common descent . Its just data.
Common design on creation week would also do this.
Your fighting a creator but rejecting a creator could have a intelligent plan for biology like in physics. Your eliminating any other options at the gate.
there is no evidence for mammals being related in a common descent way.
All there is IS like traits.
Eyeballs alr alike because of a eyeball blueprint. A very good idea. What I would do.
your saying a creator , to prove he created, must have every creatures eyebals different in structure and dna OTHERWISE you are forced to a conclusion eyeballs are the same because of common descent.
Creationists win this argument and will win it.
Thats why these ID authors win such reader audiences from people not normally interested or on the creationist side.
They really do make a damn good case and the opposition, on many points, flops.
Well lets see if anyone hear can answer my criticisms of Harshman’s criticisms.
True, it wasn’t quite clear to me that it’s congruence only when it’s the same congruence observed across several species. And why was it not clear to me? Because your definitional passage was not making it clear. Yet I understood that congruence should be observed across several species to really be congruence and this is the point I was clarifying.
Now, this clarified, namely that congruence is a set of homologies that must be present in descendant species and in the ancestor, what prevents you from saying that the descendants are really inheriting the congruence, not just a homology? Because a single ancestor can’t have congruence? A single taxon cannot either, but we already have more taxa with congruence and because of the congruence they are grouped together in the tree, thus the congruence is their common inheritance. No?
Erik,
Think ‘congruence’ in the geometric sense, as it relates to the putative tree of relationship. A real tree of descent would have a particular shape – branching order, branch length. Two trees with the same basic structure would be geometrically congruent. Of course, you don’t have the real tree. You’re trying to infer its shape from what you have. But if different characters assemble into trees of the same basic shape – are congruent – you increase confidence that there is homology – real commonality through descent.
This is not helping. Congruence in geometric terms means perfectly overlapping, basically identical in normal usage.
Then there is “similarity” in geometric sense. E.g. all circles, big and small, are “similar” and those that have equal size are also “congruent”.
When Harshman says “Mammals, for example, […] all have hair, mammary glands, and other more obscure characteristics like seven neckbones and three earbones.” and at the same time it’s common knowledge that e.g. the seven neckbones in dogs are not of the same size as in giraffes, then is this congruence in biology? Is this similarity (in geometric sense) all there is to it or is there some added nuance to biological congruence?
ETA: Another wild guess – “congruence” refers to the trees built on different categories of evidence. E.g. first you take the morphological data from taxa and you build a tree. Then you take the genetic data and you build another tree. Both trees turn out connecting the taxa in the same ways and this is called congruence. Right?
Pretty much. But congruence comes in degrees though, because you could have a tree that matches on 10 out of 12 branches, and that tree would be more congruent than a tree that only matches on 3 out of 12 branches.
As I understand it trees can go anywhere from being highly incongruent in the sense that they don’t at all lend any support to a genealogical relationship, through a sort of grey zone where the results are just too uncertain to point one way or the other, to a match that would be perfectly congruent.
Erik,
So you are incapable of using that as a starting point to understand the less strictly precise congruence of tree topologies? Come on, man. It really is like feeding a kid.
No. You still don’t seem to understand what “congruence” means here. You think it refers to the similarity of a single character among species. Actually, it refers to the same pattern of presence or absence in multiple characters among species. It isn’t that 7 neck vertebrae in dogs is congruent to 7 neck vertebrae in giraffes. That isn’t what I’m talking about at all. It’s that the character “number of vertebrae” and the other character “number of ear bones” have a congruent distribution, i.e. throughout all mammals. It’s the relationship among different characters, not the same character in different species.
I don’t know what you mean by “inheriting a congruence”. If X is inherited from a common ancestor, X is a homology. If not, it isn’t. That’s all.
Careful there when undermining Harshman’s case. It might collapse.
If “trees can go anywhere”, then “true homologies” (Harshman’s term) might not mean what you think they mean, provided that they exist at all.
I said, and you quoted from me, “congruence is a set of homologies”. By now you are reading misconceptions into what I wrote. Perhaps deliberately.
I understood this very well. And this means Rumraket has not understood this. On another note, it might have helped if you had defined “congruence” like this up there in the OP.
Understood. And also, if it is not congruent, then it’s not a homology. Right? Certainly not a “true homology”.
Careful, you’re starting to get paranoid. When you started talking about a single character, 7 neck vertebrae, as being “a congruence”, that’s when I became sure you weren’t understanding.
If what isn’t congruent? And congruent to what? If two characters don’t fit the same tree, then at least one of them is not a homology. Perhaps this would be simpler with A, C, G, and T rather than neck bones. Say we have four species, W, X, Y, and Z. If at site #1, W and X have A while Y and Z have T, and at site #2, W and X have C while Y and Z have G, that’s evidence that the bases at the two sites are homologous. If, however, at site #3 W and Z have A while X and Y have G, that’s evidence, in the context of the other two sites, that the bases at site #3 are not homologous. Is that clearer?
In your most recent example (just prior to where I am quoting from), X. Congruent to Y, presumably.
Not at all, because now you are talking about something called “site”. What is that? The location of a population or the location in the anatomy where you are examining the characters? Something else?
Never a good idea to keep bringing in more and more new terms.
No. X and Y are species. Species aren’t congruent. Characters are congruent. In the example, the pattern of bases at site #1 is congruent to the pattern of bases at site #2 but incongruent to the pattern of bases at site #3.
Let’s keep trying. For further clarity, I change the letters for species to lowercase.
Site #1: w and x have A; y and z have T
Site #2: w and x have C; y and z have G
What exactly is supposed to be homologous to what here? The species? Because they are only things that line up.
Homology was supposed to be “similarity due to common ancestry” but this scheme displays zero similarity. The uppercase letters now cannot correspond to earlier neckbones or characters or whatever.
And what the hell are “bases”?
Sorry, I thought you would have understood that A, C, G, and T are the four bases of DNA. A site is a particular spot in a DNA sequence. If we line up the example, there are four (very short) sequences that look like this:
W ACA
X ACG
Y TGG
Z TGA
See how the first two sites have the same pattern while the third has a different one?
So you mean
species=site1…site2…site3
W=A…C…A
X=A…C…G
Y=T…G…G
Z=T…G…A
“Pattern” meaning that species w and x are to be grouped together by the first two sites, but they cannot be grouped together by the third site?
Yes, I see now. But this is as unimpressive as it gets. If genes consist of letters like this – and just three letters – then of course they sometimes line up and sometimes don’t. How they came to be arranged like this (as in common descent or not) seems to be a totally different issue, nothing to do with the lineup.
Neckbones and other characters were a much clearer example.
But of course genes don’t consist of just three letters. They consist of thousands of letters. Two species would line up at one site, purely by chance, 1/4 of the time. Multiple species would all line up by chance much less often. If they line up more than that, it demands an explanation. Common descent is the explanation. Nobody has been able to think of another one.
Going back to the example, we would provisionally consider the two As at site #1 in species W and X to be a homology and infer that there was a common ancestor of W and X (more recent than the common ancestor of W, X, Y, and Z) that also had A at site #1. The more sites that match that pattern, the more we believe the homology.
From my point of view, this is completely upside down. I don’t think anything in nature, certainly not in living organisms, is due to chance. So all similarities and lineups make perfect sense and they demand no explanation whatsoever. What demands explanation and seems worth studying is the range, mechanisms and causes of variation.
Wow, Erik, you’re in for a treat if you make the effort to find out!
I doubt you can defend the claim that nothing is due to chance. A lot of things sure look like chance. But of course the similarities in DNA sequences aren’t due to chance. They’re due to inheritance. You can’t just say they need no explanation. What is your explanation, if not inheritance?
Within species, yes.
How do you determine this across species?
Simple enough. By the standard method of science, which is to ask whether one hypothesis explains the data much better than some other hypothesis. If the data show a strongly nested hierarchical pattern, we infer that the similarities are due to common descent, because we know that common descent would be expected to produce such a pattern and we can’t think of anything else that would be expected to do so. Do you?
Dead end. Hypotheses are ridiculously easy. Proving them is the hard part.
Astonishing lack of imagination. Basically, you prove your hypothesis by failing to think of any better hypothesis.
Wells is wrong in asserting overt circularity in the concept of homology. The problem with the concept of homology is something different: Homology, on the current flimsy basis, is not evidence.
I can dream up tons of hypotheses. This is beside the point. What matters is the relevant analysis and conclusion. Currently, in purely scientific terms, the evidence is inconclusive for common descent. Yes, there are similarities and then there are closer similarities. But none of the similarities deserve to be called “due to common descent” at this stage.
Shared accidental features. What Harshman says, of course, but the accidental features existing across species are especially good markers, like the accidental features of shoeprints, fingerprints, or plagiarized writings.
Almost any gene could be significantly different in its DNA, with little or no effect, yet closely related species are usually going to have the same or almost the same DNA in most of the positions of its genes. That’s not because of function, which is served equally well by substantially changed DNA in rather more distantly-related species, it’s just because they share a fairly recent common ancestor. Same for ERVs and other DNA sequences that lack significant function.
Accidental characteristics without any significant function are generally understood as revealing a certain kind of origin that would cause those accidental characteristics, from crime solving to phylogenetics. Life is full of such accidental features.
Glen Davidson
So your basic problem is that you have no idea of how science works. All science works the way I have just described. You’re just rejecting the whole process of science. Now, if you really can dream up tons of hypotheses to explain the nested hierarchy of life, why not dream up just one right now, and we’ll see whether it explains the data better than common descent? Bet it doesn’t. But that’s how science is done.
I know how science works. It’s just that different sciences operate with different standards of evidence.
Then something like common descent should be the standard in any science where you can draw trees.
They are just similarities. You can draw nested hierarchies in linguistics too. For Darwin, this analogy looked like a very compelling confirmation for common descent. Except his assumption that all languages derive from a single one was false… Well, maybe it wasn’t. The standard of evidence in linguistics forces the conclusion: The evidence is inconclusive.
I don’t understand this comment. There are number of natural and a number of man-made items that can be displayed in tree configuration relationships: products of erosion, tornado formation, the history of cars, the history of buildings, and so forth… Yet it would make absolutely no sense to infer common descent for such phenomena for one simple reason: none of those things reproduce and pass on defining information.
Similarly, I don’t understand how anyone can take similarities across species as evidence for common descent. Don’t you see the differences or what?
The nested hierarchies in linguistics are evidence that the languages in the hierarchy are related. And the standard really are the same. There is no doubt that English is a Germanic language and that Germanic languages are part of the Indo-European group. (And the presence of linguistic fossils, i.e. ancient documents, doesn’t change the method. Would you also deny Algonquian, for which there are no such fossils?) That the evidence is less conclusive for the connections among IE and other language groups is irrelevant to Darwin’s point, which wasn’t about universal common descent, just about how one can infer common descent. Your harping on the universal common descent of all language is irrelevant and has nothing to do with Darwin’s analogy.
Now, if you wanted to argue about Nostratic, the question wouldn’t be whether you can draw a tree, or how it would be interpreted — it would be a hypothesis of descent — but about the quality of the particular data used to relate, say, IE and Uralic-Altaic. The inference process is pretty much the same.
But I wish you would stop harping on that trivial analogy.
Why trivial? Is there something about linguistics that gives you a rational reason to say so or is it just that you don’t like the analogy? Yet Darwin made this analogy.
Perhaps you know a Nostraticist who has combined all the language families of the world so as to be as scientific as biologists are? Because that would be common descent in the relevant sense. The obvious families like Germanic etc. are, should I say, trivial, particularly given the way we know directly, observably, how language changes. We know nothing like this about species. Yet Darwin thought he was making a good analogy.
Because it was not intended to carry the weight of Darwin’s argument, as you seem to think. It was just a passing attempt at clarification.
No, but I know of Nostratic, which includes a number of language families. Once again, you’re looking for an analogy to universal common descent, which is not the subject. There’s a lot of room between separate creation of every species and common descent of all species, and that’s the room Darwin was occupying, and it’s also what we’re discussing here. Most phylogenetic analyses do not attempt to relate all the species in the world, just subsets. That’s analogous to relating Indo-European languages, or even the Nostratic languages. Darwin, in the origin, didn’t even mention universal common descent. The closest he got was “…breathed into a few forms or into one”. Your obsession with universality is pointless and ahistorical.
It’s because of variation within species resulting from reproduction and mutation as well as the gradient of differences across species that supports the inference of common descent. If organisms weren’t actually related, the differences between them would be much more exaggerated than what we see simply because the variations within species would be much more distinct from the variations in other similar species. But we don’t see that.
He didn’t mention this exact string of words, but he meant the same thing by various phrases, e.g. “community of descent”. Emphasis mine,
He doesn’t say “In the beginning there was just one cell/species” but that’s the logical conclusion of his trail of thought, because no matter what you compare, you will find some similarity. If this passage is any guide, where there’s a similarity, there must be a grouping.