
This is the third and last of my answers to Jonathan Wells’ 10 unanswerable questions for evolutionists.
Question 3: Homology.
Wells: Why do textbooks define homology as similarity due to common ancestry, then claim that it is evidence for common ancestry — a circular argument masquerading as scientific evidence?
This question stems from confusion on Wells’ part between how something is defined and how it is recognized, which are two quite different things. Homology is indeed defined as similarity due to common ancestry. But we don’t just label any similarity a homology and call it evidence for common ancestry. That would indeed be circular. What we really do is quite different. Similarity between the characteristics of two organisms is an observation. If the similarity is sufficiently detailed (“both are big” or “both are green” won’t do) we consider it a candidate for homology.
Homologies can be tested to some degree by predicting that the characters will be similar in ways we haven’t yet checked. For example, if we propose that similar-looking bones in two animals are homologous, we might predict that they would arise from similar precursors in the embryo, have similar spatial relationships to other bones in the organism, and have their development influenced by similar genes. And this is commonly the case.
But the main way of testing candidate homologies is by congruence with other proposed homologies. By congruence we mean that the two characters can plausibly belong to the same history. If the history of life looks like a tree, with species related by branching from common ancestors, then all true homologies should fit that tree; that is, each homology should arise once and only once on the tree. If a large number of functionally and genetically independent candidate homologies fit the same evolutionary tree, we can infer both that the candidates really are homologies and that the tree reflects a real evolutionary history.
And in fact that’s what we commonly find. Mammals, for example, are inferred to descend from a common ancestor because they all have hair, mammary glands, and other more obscure characteristics like seven neckbones and three earbones. All these characteristics go together: mammals have all of them and no other animals have any of them. Further, other characters support consistent groupings within mammals, and groupings within those groupings. Within most of life, groups are organized in a very special way called a nested hierarchy. In a nested hierarchy, every group is related to every other group in one of two ways: either one group entirely contained within the other (as in a below), or they share no members at all (as in b below). No two groups can partially overlap (as in c below).
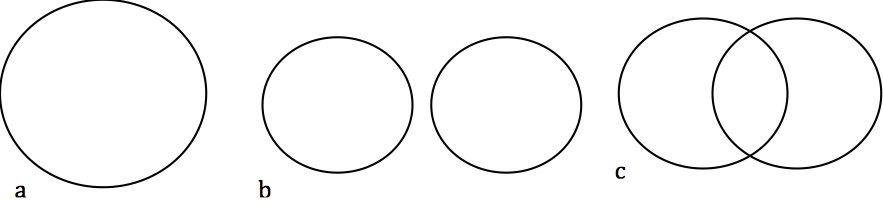
What we see if we try to organize species using candidate homologies is that groups organized according to different characters fit together like a and b, but not c, so we get a pattern like this:
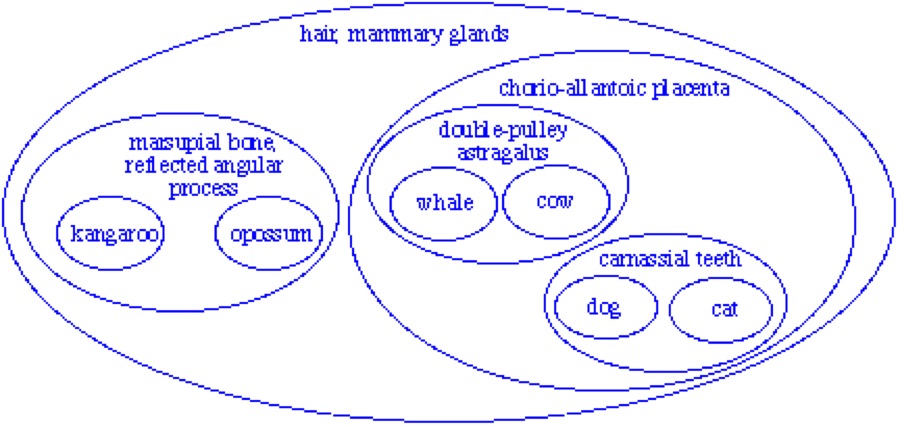
Why should these and many other characters all go together in this consistent way? Evolutionary biology explains these characters as homologies, all evolved on a single tree of descent, like this:
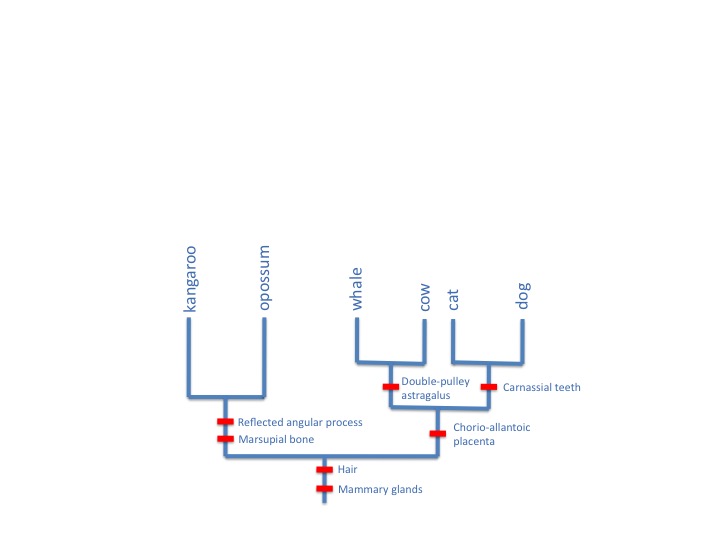
Wells gives no alternative explanation for such patterns, and indeed they are hard to explain in any other way than as reflections of an evolutionary history. Wells has it all wrong. Homology isn’t a circular argument, it’s a branching tree of evidence.
Erik,
Let me suggest that you are completely failing to understand what Darwin meant. I’ve shown you what he meant, quoting from the Origin itself. And why is what Darwin meant even relevant to modern evolutionary biology? He’s not the seal of the prophets or anything like that. How is this at all relevant to what homology is and whether we can infer phylogeny?
Deal.
(Because to bury the originator of the modern evolutionary biology is to bury the modern evolutionary biology. If even Darwin didn’t get evolution right, then nobody can.)
I’m afraid you’re treating Darwin as if he were the seal of the prophets. He was wrong about a number of things. We know much more than he did. Science progresses, you know. So what you’re saying is absurd.
And he was actually right on the point we’re talking about now, though you seem unable to see that.
John Harshman,
Thats a error. Yes German brought forth english etc but no indo european did not bring forth german.
Or rather the evidence for it is entirely based on likeness in words etc.
Instead there was one language, at Babel, and it was just a sudden twist on big identity groups to be different from others. No evolution.
no reason to see a indo european language.
the reasons for why they are related does not demand common decent mechanism as it does in a german family relative to english.
Its just reasoning from here back to there without allowing other options.
Erik, observe your doppelganger, differing only in that he’s ignorant of linguistics as well as biology.
When a species gives birth to another species, what exactly is it that corresponds to the zygote in that process?
LoL!
It’s like a shell game. Follow the pea.
John’s playing his games again. Trying to shift the burden of proof. Some things just never change. So we have hypothesis J (the one John likes), and John and cohorts have compared it against what other hypotheses, exactly?
We know that the “science” of evolutionary biology doesn’t work like the science of physics. So we seem to be a step ahead.
Wells, Jonathan. Zombie Science: More Icons of Evolution (Kindle Locations 310-311). Discovery Institute Press. Kindle Edition.
Wells, Jonathan. Zombie Science: More Icons of Evolution (Kindle Locations 314-316). Discovery Institute Press. Kindle Edition.
Wells, Jonathan. Zombie Science: More Icons of Evolution (Kindle Locations 322-326). Discovery Institute Press. Kindle Edition.
Wells, Jonathan. Zombie Science: More Icons of Evolution (Kindle Locations 425-428). Discovery Institute Press. Kindle Edition.
Really, wow…there is a way “science” works, cool!
Political science? Sports science? Science of sleep? Science of hypnosis? The science of love?
Please tell me again where you described this, this is amazing news. The science of science!
Disciples of Darwin are often stuck in the 19th century.
Sir Isaac Newton got some things wrong
Was gravity among those things?
He predicted the effects correctly for the most part without knowing exactly how it worked.
Mung,
When a bacterium, through fission, results in two descendant bacteria, what exactly corresponds to the zygote in that process?
Absolutely nothing. The ball’s back in your court.
Wells, Jonathan. Zombie Science: More Icons of Evolution (Kindle Locations 451-453). Discovery Institute Press. Kindle Edition.
Wells, Jonathan. Zombie Science: More Icons of Evolution (Kindle Locations 466-468). Discovery Institute Press. Kindle Edition.
doplleganger seems a foreign word , never heard it before, but i’m not ignorant of linguistics, or its not revealed by why i revealed about this talk on linguistics.
The bible said all spoke the same language. At babel everyone was divided into family groups. then god confused, put a spin, on the language.
Just a spin. not whole new languages.
so it would be, if this was so, that all languages could show likeness.
Yet the indo -european languages did not evolve from a common original language but are only twists off the great parent language. So the indo-eur group just reflects a closer group. its the Japhet group. A son of noah and shown in the table of nations in genesis.
Again this is a classic case for insisting there is only one option.
BECAUSE there are other options for likeness, other hen common descent, then subjects like homology are wrong to define homology as from common descent.
Then use likeness to say they proved common descent like Wells smartly pointed out.
Homology is only about likeness and not about why its likeness.
Please correct the textbooks from the evolution bias and incompetent scholarship.
Homology back to the people.
Mung,
What? It just whizzed off the edge of your racket into the bushes. You think that the only sense in which something can have ancestors or descendants is by mating and forming a zygote? That’s silly. New balls please.
Why don’t you start the ball rolling by showing what such a correction would look like. Why don’t you write chapter one of the new textbooks? Or is it that someone else, as usual for you lot, has to do the actual work?
The trouble with that is, he just might do it. Not at all well, of course, but he has written a fair amount about how marsupials are merely pouched versions of placentals. Even creationists won’t swallow it, but he seems never to doubt himself, no matter how wrong.
He won’t do the science, of course (doesn’t understand it), but I wouldn’t encourage him to write nonsense. He can do that.
Glen Davidson
i’m the thinker. writing is another skill.
Anyways i’m making a excellent case for why everyone should change the textbooks.
they are clearly wrong on homology. Well’s is completly right and is writing about it and maybe will be a agent of change in biology textbooks.
Homology is just about bodyplans/traits looking alike in different creatures.
So why is the definition of homology based on common descent??
A definition of homology must be independent of any hypothesis of its origin.
This is the law of science definition!!
Then to rub salt in the wound.
They use , in contention on origins of homology, the definition as proof for the claim of homology coming from common descent. oh brother.
Maybe i should write the biology books..!!?
Creationists aren’t right about everything?!
I don’t doubt I’m right but its from analysis and study.(actually lots of humanity don’t doubt themselves and is a problem)
I understand the “science concept”. Its just people thinking and doing a better or worse job.
By the way marsupial convergent evolutionists claims relative to placentals is a issue for homology based on common descent.
Body plans/traits in this case contradict what they do otherwise.
.
“what they do otherwise” suggests that all (or most) non-marsupial mammals provide body plans/traits which fit well into common descent homology. I would strongly agree with you on this point if no other;-)
And if Comparative Linguists starting considering “likeness in words” between languages as objective evidence, where would we be?