
Since I’ve been asked, I’m posting another of my answers to the 10 unanswerable questions for evolutionists in Jonathan Wells’ book Icons of Evolution.
Question 5: Archaeopteryx.
Why do textbooks portray this fossil as the missing link between dinosaurs and modern birds — even though modern birds are probably not descended from it, and its supposed ancestors do not appear until millions of years after it?
What does Archaeopteryx have to be to qualify as a “link” (not a missing link, because it isn’t missing)? Wells apparently (he never really says) requires an insensible gradation of ancestors and descendants leading from an unquestioned dinosaur to an unquestioned bird, with Archaeopteryx in the middle. While that would be nice, it’s hardly necessary — and considering the quality of the fossil record, that’s lucky.
How likely are we to find direct ancestors of living species in the fossil record? That depends on the quality of the fossil record. If we have found most of the extinct species that ever lived, our chances are good; on the other hand, if our knowledge is spotty, our chances are bad. We can judge the quality of the dinosaur fossil record based on the species we have found so far. Half of all known dinosaur genera are known only from a single specimen, which suggests that there are many more genera for which not even that single specimen has yet been found. Moreover, many of the genera with multiple specimens are known only from a single time and place. We have seven specimens of Archaeopteryx, all from a single limestone quarry in Germany. Archaeopteryx is the only known Jurassic bird. How likely is it that the single Jurassic bird we happen to have found is the ancestor of all subsequent birds? Given the small sample we have, we are unlikely to have found the ancestors for most dinosaur groups, including birds. Fortunately, we often can find fossils that are not too far removed in time and appearance from those ancestors. Archaeopteryx is one such fossil. It probably isn’t the ancestor of birds. (Distinguishing actual ancestors from cousins of the ancestors is itself an unsolved problem.) But it does represent a key transitional stage between theropod dinosaurs and modern birds. It has some features of theropods, some of birds, and others that are in between. Wells offers no explanation for the transitional nature of Archaeopteryx. We have other fossils for both more primitive and more advanced transitional stages. Some of the more primitive transitional stages — Velociraptor, for example — lived later than Archaeopteryx. Such are the vagaries of preservation. Nobody claims that ancestors appeared after their descendants, only that we have sampled a big family of cousins and siblings at random points through time, some of whom (like Velociraptor) resemble their common ancestor more closely than others (like Archaeopteryx).
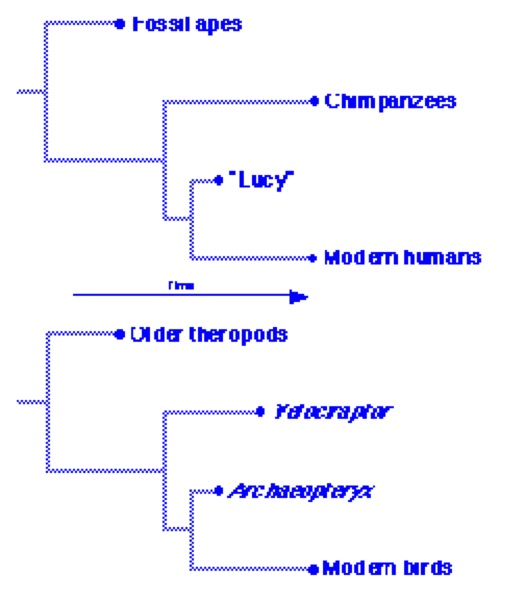
A comparison may be helpful in understanding this point. On the top of the figure above is a tree showing human relationships. Dots at some points in time represent species we know about. We have a number of fossil apes, most over 10 million years in age. We also have a number of fossil hominids, of which Australopithecus afarensis, represented by “Lucy”, is perhaps the most famous and has often been mentioned as a possible human ancestor. While it’s not clear whether A. afarensis is or isn’t a direct human ancestor, it’s definitely not too far from that line. The next closest human relatives are the chimpanzees. Chimpanzees have no known fossil record. Someone might try to cast doubt on this human phylogeny by claiming that modern humans are probably not descended from Lucy, and her supposed ancestors (meaning chimpanzees) do not appear until millions of years later. Of course, there are other, earlier fossil apes, but they are much farther from humans than are chimpanzees. Nobody actually makes this argument, probably because we easily recognize it as ridiculous, equivalent to the question, “If humans are descended from apes, why are there still apes?” You don’t even have to accept human evolution to realize that question is silly. But Wells’ question about Archaeopteryx is exactly the same. We’re not sure if Archaeopteryx is a direct ancestor of modern birds, but it’s not far from that line. The theropod dinosaurs that Wells calls Archaeopteryx’s “supposed ancestors”, like Velociraptor, are merely the closest known relatives of Archaeopteryx, and stand in the same relationship to it that chimpanzees do to Lucy: later in time yet more primitive. And of course there are plenty of earlier, even more primitive theropod relatives. We know that the fossil record is incomplete, and it’s incomplete for both birds/dinosaurs and humans/apes. The fit between the actual fossil record and our natural expectations that primitive characters will appear earlier than advanced characters is surprisingly good, but it’s not perfect; as in these two cases, some fossils we would like to see have not been found. But enough have been found to give us a clear picture, at least in outline, of both human and bird evolution. “Lucy” and Archaeopteryx don’t have to be directly ancestral to make important contributions to those pictures.
OMagain,
They use the same design concepts that are used in semiconductor designed communication systems. A translatable code and repair mechanisms are examples.
How would you support the claim that this is the result of a blind process?
Allan Miller,
Where is the evidence that supports the long term mutation rate? If it is real why wouldn’t we see it in the DNA sequences?
Allan Miller,
With all due respect you have not supported this and if you read up on DNA repair you will see that your claim is incomplete.
colewd,
Here’s a longer answer:
1) polymerase error repair detects the ‘lumpiness’ that occurs when a base is not in a Watson-Crick pairing A-T or C-G. This depends only on information in the pairing – the two strands. Unless you know different.
2) Homologous repair pathways (there are several) depend generally upon the information in the homologous chromosome in a diploid cell. Not all of them are non-mutagenic, contrary to your supposition. In fact NHEJ (nonhomologous end joining) is the commonest repair pathway through much of the cell cycle, and that doesn’t look at the homologue at all, rather surprisingly. It is mutagenic. But, it doesn’t get any info from outside the immediate genome either way. Unless you know different.
3). Apoptosis pathways in the germline can only work by gross reference to the genetic complement of ‘normal’ cells, or distinctive features of ‘broken’ ones. It makes no sense to go outside the individual, since every normal cell in the population would appear in error against that standard, given recombinant genomes. Unless you know different.
colewd,
With all due respect it was supported exactly as much as you supported your ‘mean’ contention. I’ve now gone further. Your turn.
Allan Miller,
The current evidence supports this claim.
The other two are not supported to my knowledge but I will do some research here. If you can support thank you in advance for saving me the time.
That is not evidence of design.
Name as many as you wish, that is not evidence of design.
The work towards supporting that as a blind process started some time ago, had a notable boost from a Mr C.Darwin and since the the evidence has only continued to accumulate. And in all that time, no sign of nor any need for a designer.
I’m asking you how you would support the claim that cells are designed and you respond by saying they use “design concepts”. Then you ask how I would support the claim that cells are the result of a blind process.
I would advise you to concentrate on working on improving the support your claim that cells are designed, because as noted since Mr Darwin kicked the ball into overdrive all the evidence so far has accumulated on one side. And it’s not the “designed” side.
Rain falling on a cracked ice sheet, freezing it over and smoothing it is presumably a repair mechanism then for the ice sheet. Therefore winter is designed?
OMagain,
I gave you examples of design concepts that are in the cell. You have not given an example of a blind process creating something that functions repeatably such as cell division.
A lot has been learned about cellular function since Darwin and the blind unguided story is clearly losing steam as we learn more about how cells function.
What does any of this have to do with Archaeopteryx?
It’s really just how it has to be, there is no other physically realistic or logically consistent option.
Just turn on your brain for a moment and you’ll see why that is. In case this is beyond your mental capacities, here it is: If for some reason an organism is born by parents that belong to some species, but the newborn organism cannot interbreed with any other contemporary member of that species, then whatever new species you might argue the newborn organism belongs to, will go extinct with that single member since there’s no other individual to breed with. The same could not be said for asexual organisms of course. So IF macroevolution is true, it HAS to be the case that all new members born at any particular point in time will be of the same species as it’s parents.
No, you merely claimed with zero evidence that they are “design concepts”. They’re not “exampels” of design concepts just because you desire intensely to think about them in that way.
So now we’re supposed to show you how the very first cell originated, otherwise you’re just right by default, is that how this works?
A lot has been learned about cellular function since Darwin and the blind unguided story is clearly losing steam as we learn more about how cells function.
Yes yes bla bla bla evolution will soon be dead. ZzzzzZZz
Phylogenetics.
Now how would YOU support the claim that they were designed?
Well, now. Phylogenetics doesn’t tell you evolution was a blind process. All it tells you is that some particular group of species is related by descent in some particular way. How the mutations that unite and separate the species got to be that way isn’t something phylogenetic analysis really covers. (Though we do use models of the evolutionary process in estimating the trees, that’s not quite the same.)
John Harshman,
You are right John. Sorry for getting side tracked.
Rumraket,
As I understand it, Chromosomal speciation (e.g. Blind Subterranean Mole Rats [Spalax] and Rock Wallabies [Petrogale]) leads to offspring that may great difficulty breeding with member like their parents, but not others with a similar chromosomal rearrangement. Though the parents and offspring would seem identical functionally and morphologically, the reproductive rift is the beginning of a speciation event.
All true, but I really don’t consider the option that God caused the mutational differences we see between different alleles in different species, to be on an equal rational or evidential footing as the option that they were caused by basically the same processes of mutation we see operating in the here and now.
You could then say, I guess, that even for the mutations that happen now in genomes of extant organisms, we can’t somehow prove they’re not being caused by God. True, but why would you even believe that in the first place?
Are we really in the situation where past historical events can equally well be attributed to divine causes? I don’t think so. I think this really comes down to Occams razor. It’s not that it isn’t at least logically possible that God personally intervened for every mutational event in history, it’s that it’s simply not necessary to invoke that to explain what we see. That the sequence differences we see were caused by already well-known biochemical reasons and filtered by a blind sampling process, is a simpler explanation, than to multiply entities beyond what is needed and start including intending agents as the causes of particular individual mutations in the deep geological past.
Simpler, but far less facile.
Glen Davidson
That is interesting, and may be an exception to what I thought I understood about how speciation might generally proceed (among other things I thought it was generally accepted that geographical isolation is one of the main contributing causes, because it prevents gene-flow between members that otherwise would and could interbreed).
But I have a really hard time seeing how this could be a general mechanism of speciation. It seems to rely on the idea that these chromosome-scale mutations are something that happen frequently enough, yet are systematically similar across the population in a way that makes them reproductively compatible for individuals that have them, that a substantial enough fraction of the population at any given point in time will have offspring that can go on to sort of “seed” a new species.
This might be the case for Mole Rats and Rock Wallabies (and plants?), but I don’t believe this is normally how animals speciate.
Rumraket,
‘Absolute’ chromosomal speciation seems likely to be rare for mechanistic reasons. If it is an absolute barrier to reproduction with the parent species, there would have to be two offspring with the same mutation, of opposite sexes, for the lineage to stand any chance in an obligately sexual species. If it merely depresses the fitness of heterozygotes, it may create a lever. But there is a limited time frame during which something else has to happen to reinforce it. Eventually, one or the other pattern may still drift to fixation, despite this leaky heterozygote barrier, assisted by increasing numbers of homozygotes as it becomes more common.
Language evolution both indicates the truth that all evolution isn’t exactly dependent on “random mutation,” and the fact that invoking a directing super-intelligence really is a pathetic explanation even for language evolution. For, it’s not like brains aren’t involved in both language stasis and change, yet there’s no intelligent decision about hanging onto irregular verbs, keeping a silent “h” around like the French do (with inconsistent rules), or keeping old pluralizations around for commonly used nouns (children, men, women) in English.
Language evolution has atavisms like biologic evolution does, mainly because although intelligence isn’t exactly missing from language evolution, neither does intelligence necessarily overcome legacy, inheritance. No one would intelligently decide to change languages like they do change, even though brains are involved with language evolution.
The speed at which language evolution occurs is due to the fact that they are not dependent upon random (with respect to fitness) mutation, nor is there any biologic counterpart to broadly rule-based whole language changes, like the vowel shift that occurred in English. Where the analogy between language evolution and biologic breaks down is where the brain is involved and thus makes language far more labile and the changes far less due to randomness. Nevertheless, both sorts of evolution are indicated by the effects coming from the fact that no great intelligence directs the changes that we see, rather, conservation and change occur according to factors other than intelligence rationally controlling the outcomes (not that no rationality is involved with languages, of course, but that rationality fails to actually redesign languages to be quite rational, like Esperanto (I assume)).
I would say that evolutionary trees are quite possible without random (with respect to fitness) change, but they are not consistent with some great intelligence rationally designing either languages or life to make them as consistent and reasoned-out as they can be. One has to be as cognizant of where analogies break down, though, as where they do not, and realize that while biology isn’t dictated by some great intelligence, neither is it normally controlled by brains at all, so that there is no rational rule-based change occurring throughout life (or even a species) as there can be throughout languages. Changes have to be compatible with the rest of the organism (and its interactions with others, to be sure) to be kept, and especially, in order to become fixed, but they don’t have to agree with some mental sense of consistency with both the past and, say, shifting conjugation rules (or not, as the case might be). Intelligence doesn’t rationalize either language or life, but brains are involved with the former and (apparently) not the latter.
Random (with respect to fitness) change is all that life gets for new information, according to the evidence, so to be consistent with the evidence it won’t do to invoke intelligence even at the lower level, as one has to with language evolution. Some evolution that is somewhat analogous to biologic evolution is possible having brains in control (but without overall planning), however, brains do not control anything (other than, like, mate selection, in a few species of organisms) with respect to biologic evolution, so far as we can see.
Glen Davidson
Agreed. It’s just that phylogenetics isn’t what tells us that.