
I am sure that many readers have already concluded that I do not understand the role of sex in either organic or biotic evolution. At least I can claim, on the basis of the conflicting views in the recent literature, the consolation of abundant company.
– George C. Williams, Sex and Evolution, 1975
What’s sex all about? This question has been exercising biologists since well before Williams’s time, but in the 1970’s, with the rise of ‘gene-centrism’ and the related controversy over group selection, a succession of prominent authors grappled with the problem, trying to fit it with current evolutionary theory to no-one’s particular satisfaction. Males were deemed an impediment to a female’s efforts to maximise her reproductive output, time wasted on these feckless types resulting in her only passing on 50% of her genes per offspring. From the perspective of a ‘selfish gene’, meanwhile, getting into every offspring seems a preferable fate to only getting into half of them. On the basis of these apparent large costs, a cryptic offsetting benefit of corresponding magnitude was assumed. Like Godot, it is yet to appear.
Yet sex is widespread. All eukaryotes either do it now, or possess tell-tale signs that their recent ancestors did. Given that it appears costly to individuals, and genes, how did it evolve and why does it persist?
Terminology.
In asexual reproduction the entire genetic complement of cells or individuals – all of their chromosomes – is duplicated during mitosis. Two new cells are produced per complete division, giving exponential increase where unrestrained.
Mitosis occurs in sexual lineages too, but the defining characteristic of eukaryotic sexual reproduction is a cyclic alternation of ‘ploidy’, or chromosome number, interrupting the mitotic series. A haploid cell has one set of chromosomes, a diploid two, and so on, and in sex, two haploid gametes merge to form a diploid zygote in which each chromosome has a similar partner – a homologue – in the other haploid set.
At some future point, the reduction division of meiosis generates more haploid sets from the diploid, by pairing then segregating these homologous copies into new haploid cells. Additionally, during haploid separation there may be reciprocal swap of entire chromosomes or segments: recombination. By this process, previously linked genes end up in different individuals, while previously separate genes end up linked.
While in the diploid state, at any segment held in common between a pair of homologous chromosomes, the genetic sequence may be identical (homozygous) or not (heterozygous). Where heterozygous, the genome has two different alleles (sequence variants) at that locus, one of which may be dominant (expressed) and the other recessive (suppressed).
The problem
Consider an organism … with equal numbers of males and females … In females, a gene A suppresses meiosis, and causes the production of diploid eggs that develop without fertilization into females genetically identical to the parent. … when rare, such a gene would double in frequency in each generation. This result has been expressed by saying that there is a ‘twofold cost of sex’, arising from the needless production of males. It is clearer, however, to take a ‘gene’s eye view’: a gene A that suppresses meiosis is certain to be transmitted to all the eggs produced by a female, whereas a gene a that permits meiosis is transmitted to only half.
John Maynard Smith, Evolutionary Genetics 2nd Ed 1998.
The issue tends to be framed in population-genetic terms, by reduction to mathematical abstractions or computer models based on the diploid phase. When applying such simple models, the following drawbacks appear, when comparing sex to perpetual asexuality:
- Sex breaks up adaptive gene combinations (costs of recombination/segregation).
- Sex halves the genetic contribution of an individual to the next generation (‘twofold cost’ of meiosis).
- Sex halves the chance of a given allele getting into the next generation (cost of meiosis, gene’s eye view).
- For an organism with separate genders, sex may halve the number of grandchildren produced (twofold cost of males).
- The need to locate a mate.
- Time costs.
Sex does have accepted benefits. As Fisher and Muller noted in the 1930’s, sex, largely through recombination, can enable populations to concentrate beneficial alleles and purge detrimental ones independently, and to generate beneficial combined genotypes more rapidly than serial mutation in an asexual lineage. Sexual populations also tend to have more standing variation, which can assist in withstanding environmental change, disease, or parasites. These benefits, however, are at population level. This kind of ‘group selective’, good-of-the-species idea has a problem: how do you get from low frequency to become common enough to reap the group benefit? Especially when, on cost-driven thinking, individuals (or, if you’re a gene-centrist, genes) suffer such a high cost?
The solution (!)
My heretical contention is that the costs are largely illusory: an artefact of perspective and model. Sex doesn’t start with the diploid chicken, but the haploid egg. Regarding diploid somas as the central entity in biology is natural enough. It’s what we are, and is how most eukaryotic organisms spend most of their life cycles, and therefore forms the basis of much of population genetics itself. But in the matter of sex, it’s the wrong start point. Choosing to start with diploidy begs an erroneous view of the transaction. We will never find that elusive benefit for the diploid; there isn’t one. I argue that the entity for whose benefit sex exists is not the diploid organism at all, nor individual genetic loci, but the haploid genomes that nowadays slip almost unnoticed from instance to instance of the larger bodies they often form in diploid partnership, now thoroughly shuffled by the transaction. One might say that haploids don’t exist for the propagation of diploids; it is the other way round.
If we start with fusing haploids, the diploid cell they form is a temporary union, no more an indivisible unit with superior ‘interests’ than a human couple. Despite the superficial elaborations and apparent asymmetries in modern organisms, genetically sex remains a symmetric and mutually beneficial cycle of haploid fusion and division, and looks much the same to haploids as it always did. Diploidy is haploid symbiosis: a marriage of convenience.
While such diploid unions can and do become permanent, they inevitably appear in a background of an established sexual competitor and ecosystem; their repetitive genomes are rarely up to the challenge, while tending to suffer genetic degradation if their paired haploid genomes spend too long in harness. The tools of population genetics, with their inevitable simplifications, tend to obscure rather than illuminate the respective dynamics of the two modes – they overestimate the frequency of asexuals’ occurrence, and over-count their relative production.
The argument should hopefully become clearer if we follow the trajectory of sex from its probable start point: haploid fusion.
The most likely evolutionary sequence is:
- Haploid fusion – sex originated among haploids, not diploids
- Division – return to the haploid state via the ‘back end’ of mitosis
- Independent segregation – gives primitive, coarse recombination
- Speciation – steadily broadens the sexual clade and ecosystem
- Crossover – helps tensioning in meiosis, with far-reaching side-effects
- Multicellularity – nurtures and amplifies paired haploid genomes
- Gender – gamete asymmetry, only possible in multicellular forms.
To start at the very beginning …
1) Haploid Fusion.
The ancestral population in which sex arose, more ancient even than the most recent common ancestor of all modern eukaryotes (being itself sexual), must have been haploid. To get two similar chromosome sets in the first place, they must be copies of a single template in their most recent common ancestor, since duplicated through mitosis in separate haploid lineages. Thus, immediately we can see the that the ‘haploid-centric’ perspective is fundamental, and not merely arbitrary. Diploidy is more usually chosen as the starting point only for reasons of convention and a ‘diplocentric’ bias, rather than evolutionary logic.
There has to be a reason why such fused-haploid cells could prosper of course, but we don’t have to explain everything at once. The following are suggested possibilities for selective advantage, not mutually exclusive:
- Fusion generates an immediate increase in unit size, without going to the trouble of conventional growth. Whether predator or prey, this has the clear potential to be advantageous, to both partners equally.
- It is commonly observed that hybrids exhibit ‘vigour’, being frequently more robust than either parent even if sterile. The cause is related to heterozygosity – deleterious recessives can be masked by their dominant allele, the heterozygote may be ‘fitter’ than either homozygote, and nonoverlapping loci can ‘complement’ each other. There is no reason to suppose that this phenomenon is recent.
2) Division.
Of course, we have an immediate problem – sex as defined is a cycle, alternating haploidy and diploidy. Whatever benefits derive from fusion are discarded on division, yet without a return to the haploid stage, it’s not a sexual system. However, it is not essential that we provide a positive advantage to division. For example:
- Having fused, the cell has been pushed rapidly along its growth phase. If one recalls high school biology, Interphase, where growth and replication take place, is divided into G1, S and G2. Chromosome copies are made during S, so it is certainly possible that a fusion diploid would resemble, to the machinery of mitosis, a normal asexual cell at G2. The cell has grown, and contains approximate chromosome duplicates. Rather than needing a rationale to trigger separation, an early difficulty may have been actually to defer this automated cell division to preserve any benefits of diploidy.
- It is not a given that such early eukaryotes could actually perform mitosis in the diploid state. If they couldn’t fully mitose as diploids, then however beneficial that diploid state may have been, it could only be temporary if such a lineage were to persist.
- Genomes in diploids can suffer attrition from gene conversion events. These occur during recombinational repair, in which one haploid chromosome provides a ‘patch’ to fix breaks in its homologue. In doing so, that region can become homozygous, if it wasn’t already, potentially exposing deleterious recessive alleles to selection, or reducing any benefit derived from complementation at nonoverlapping loci. Additionally these ‘masks’ can themselves increase in number, due to further mutation. Thus, a lineage of haploids which fuses but then never divides may lack evolutionary staying power, ‘selecting for’ the capacity of division as avoidance of a problem rather than exploitation of an advantage.
On both of the latter two points, even if fusion was common and division rare, surviving lineages would be biased in favour of those with the capacity of division. On all three points, no explicit advantage to division itself is suggested. From these early ‘fusers’, it may be the case that only ‘dividers’ have left descendants to the present due to this bias in lineage survival, rather than direct adaptive advantage.
______________________
Intermission 1 – The Basic Transaction.
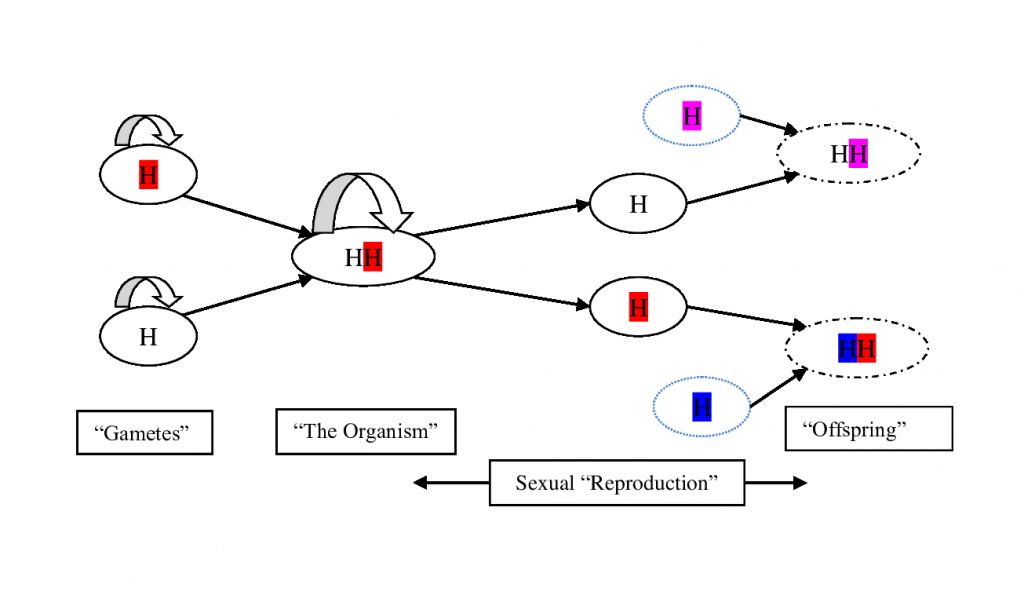
Even with this minimal system, we have all the basic components of modern sex: haploid cells fuse, form a diploid for a period and then reduce back to haploids. At this stage, diploidy need only provide a relatively minor benefit, shared by both haploid genomes, to allow it to compete in a background ecosystem of perpetually asexual haploids. Secondary asexual diploids, meanwhile, always find themselves in direct competition with a parent population already in residence, and will only be ‘average’, on average.
Let’s examine the costs in this primitive system.
Cost of meiosis
The diploid here is nothing more than a pair of haploid genomes sharing a cell, and each haploid is not obviously worse off after the transaction than another parallel pair that remained independent throughout. It is not even necessary that fusion be complete initially – the nuclei may have remained separate, as in Giardia and some fungi today. But either way, one would not view this transaction as a puzzling halving of the diploid’s genetic complement. Any benefits of diploidy accrue to both partners equally; the diploid has no say in the matter. The twofold cost of meiosis is an illusion of perspective.
The costs of meiosis and of males are often conflated. It should be clear that they are in fact distinct, despite taking the same numerical value. John Maynard Smith appears to commit precisely this error in the quote with which I headed ‘The Problem’, though he immediately points out that the cost is not applicable to isogamous organisms – those where both gametes make an equal contribution to the next generation. (Indeed, if sex really did provide a mysterious twofold benefit, as it is commonly assumed it must to cope with males, this stage of isogamy should be an absolute breeze to establish!).
Segregation load.
Segregation load is again a cost viewed from the perspective of the diploid, when comparing a population of perpetual diploids that do not return haploids to similar rivals that do. If a particular diploid combination of alleles is beneficial, sex as depicted may break that beneficial combination due to segregation. However, on this scenario it was sex that brought them together in the first place. An asexual clone will ‘freeze’ and repeat a particular combination, but it simply represents a random draw of two genomes from the wider haploid pool. The sexual population, meanwhile, continues to make random draws. There is no reason to suppose that the asexual’s luck is any better, on the average, than the sexual’s.
There is a rather puzzling assumption implicit in the genetic load arguments, that sex is fine for generating combinations, but should be abandoned the instant it has done so. Yet something better may be just around the corner – there is no reason to prefer ‘stick’ over ‘twist’. If we only count severance of adaptive combinations, without recognising the contribution of the process to their creation, we are guilty of incomplete accounting. As long as net creation exceeds net loss, sex still wins.
Variation
Gradually, as mutations accumulate, the sexual population of diploids may be expected to hold a greater degree of standing variation, even with no recombination, because its genomes circulate in ‘halves’, both uncoupling the diploid and permitting more combinations. This variation may assist local adaptation, and provide a buffer against environmental change – and, indeed, invasion by clonal asexuals.
Ongoing evolution
By circulating as, effectively, ‘half-genome’ fragments (from the diploid perspective), haploids can be tuned more readily by iteration than when locked in harness in a perpetual diploid. A beneficial mutation in the diploid state is interfered with by the other allele at its locus – if recessive, it is not even expressed. The same mutation circulating in bare haploids, however, can increase through drift, then when it begins to encounter copies of itself in diploids, can be further promoted by selection.
Gene conversion
As already noted, homologous repair can expose deleterious recessives to selection. This is an ongoing and growing problem for a perpetual diploid, which exacerbates the problems already mentioned, further diminishing the probability that secondary asexuality will extinguish sexuality. The famous rotifers – ‘an evolutionary scandal’, being a notable exception to the rule that asexual lineages are short-lived – appear to avoid this problem. Their genomes are barely distinguishable as ‘diploid’ at all. If we can’t identify homologues, the repair mechanism is unlikely to do any better. (An afterthought: that may be putting the cart before the horse. The original diploid genome may have been permitted to diverge by suppression of the use of homologues in repair).
In the sexual diploid, conversely, the negative aspects of gene conversion are diminished, since partners don’t stick together long enough for it to become an inconvenience. Additionally, it serves as an incidental mechanism of generating variation, by placing alleles into novel backgrounds.
Networking.
The sexual population of diploids is ‘networked’ by virtue of its haploid vectors. Different solutions to environmental challenges are worked on independently and combined and tested in diploids. Improved versions of the haploid genome can ultimately find themselves shared by every diploid in the future population, a luxury unavailable to the cloned asexual population except by competitive replacement of the entire species.
Asexual diploids
Of course, even at this stage there is nothing in principle to stop the diploid failing to separate, and so forming a diploid asexual lineage. However, such asexual mutants always arise within an established sexual population. A given asexual derivative has committed for better or worse to a single genome out of the myriad of possible variants available to the sexual equivalent. Armed with that single repetitive genome, we are invited to believe that, as a universal principle, this asexual genome would outcompete all variants, throughout a range, in every such contest, if sex is to retain its mystery. This seems a stretch. While the asexual might possess the fittest variant of one genotype (corresponding to a whole chromosome at this pre-crossover stage) it would be unlikely to possess the fittest variant of every single one. The resident sexual has, somewhere, an answer to every competitive challenge the clonal asexual can throw at it, it can evolve more quickly, and within it gene conversion tends to be more a blessing than a curse
In my view, secondary asexuals (those derived from an ancestrally sexual line) are better viewed as a kind of ‘species cancer’ – diploid overproduction which may eliminate the parent ‘body’ in some circumstances, but needs to do so in all to generate a ‘mystery of sex’. Cancer does not cause us to ask ‘Why People?’; likewise, secondary asexuality should not automatically lead us to ask ‘Why Sex?’. Without sex, such presumed diploid threats to it would not even exist, a neat paradox.
______________________
By such marginal degrees, then, a sexual clade may slowly rumble into life, largely untroubled, at this stage at least, by the threat of secondary asexuals, which may briefly flicker but are not up to the task required of them in preserving a ‘mystery of sex’: universal extinction of parent populations.
At what point in subsequent elaboration does the mystery commence? Does sex really have to ‘try harder’ once it discovers multicellular males?
3) Independent segregation.
Eukaryotic sexual recombination generates novel genomes by swapping segments of chromosome in the diploid. Indeed most ‘theories of sex’ are actually theories of recombination. It certainly gives rise to its most far-reaching consequences, yet the active mechanism, crossover, is complex, and its consequences seem to be at population level, requiring certain assumptions both to get it from low to high frequency, and to keep it there. Furthermore, when genetically controlled, a recombining locus may become detached from any benefit it causes.
It is not necessary to evolve something new, however. We have already touched on a mechanism by which limited recombination is achieved as a byproduct of another process: gene conversion in homologous repair. But even with this grossly simplified proto-sexual system, another form of recombination is available ‘for free’: if the haploid chromosome number exceeds 1, we see independent segregation of those multiple chromosomes, shuffling the haploid inputs. The parental haploid chromosomes are not labelled as such; the machinery of segregation simply lines pairs up at random on either side of the metaphase plate and hauls them apart. There is a 50% chance that any given former cell-mates will be separated on division: {A, B} and {a, b} ‘parents’ can produce {A,B}+{a,b} or {A,b}+{a,B} outputs with equal probability. This further increases the variation that the population can sustain.
A chromosomal break can readily drift into a population, if neutral or even if mildly deleterious. While rare, it encounters unbroken versions in meiosis, and there is no independent segregation. But when more common, it will start to encounter copies of itself, and chromosome ‘swaps’ will occur at the break point, purely by chance. As a consequence, beneficial alleles on the one are uncoupled from detrimental alleles on the other, allowing the former to increase in frequency and the latter to decrease, somewhat independently. Additionally, new beneficial combinations can arise through such swaps without the need for serial mutation in one lineage.
Note that this is precisely what Fisher and Muller proposed, but without any necessity for either elaborate mechanism or adaptive benefit to drive its fixation. Such chromosomal breaks form an ‘ideal’ recombining locus, since they do not suffer detachment from any benefit they may promote. They can drift in or out, additionally being sometimes promoted by and sometimes opposed by selection, according to the extent to which the break tends to have net negative or net positive effects. The fragments so formed, meanwhile, can increase or decrease independently of each other. The population is enriched in beneficial alleles, and similarly depleted in detriment, a comparative fine tuning not possible when alleles are chained in indivisible lumps.
______________________
Intermission 2 – On Recombination.
11112222333344445555
An idealised chromosome, each 4-character stretch representing the span of a separate gene.
Cost of recombination
Contrast the above flexible, responsive but accidental mechanism with the assumption underlying the ‘cost of recombination’: that breaking combinations is ‘always bad’. Whether it is or isn’t depends upon current circumstances. Suppose that genes 1111 and 5555 interact – in the jargon, they exhibit epistasis. This means that, in the presence of the other, each experiences either enhanced (positive) or diminished (negative) selection. If a breàk appears between them, potentially swapping 1111 and 5555 into a different background, this could be detrimental if the interaction is positive. However, if the interaction is negative, the break would be favoured.
Additionally, irrespective of interaction, genes may contribute additively to adaptation. If 2222 and 4444 are both locally adapted genes, a break between them may disrupt them, particularly in the face of a stream of less well-adapted alleles migrating in from the wider population. The combined genotype 222233334444 is assumed locally fitter than both 22223333xxxx and xxxx33334444.
In reality, though, there are thousands of genes on a chromosome, thousands times thousands of potential interactions either side of a chromosomal ‘break’, and a great reservoir of combinations to explore available in the wider population. As we saw with segregation load, recombination does not merely break combinations, it creates them in the first place. Migrants don’t merely import locally detrimental alleles; beneficial ones come in on the same tide. Thus whether a particular break point is net detrimental or net beneficial depends upon integration of a great number of variables, which the naive ‘cost of recombination’ serves only to obscure.
Selfish Genes.
The twofold cost is often portrayed from a gene’s perspective, as a halving of its chances to get into any given offspring. Richard Dawkins goes so far as to imagine genes ‘dragged kicking and screaming into the second anaphase of meiosis’ (The Extended Phenotype). His intellectual predecessor John Maynard Smith framed it less dramatically (see ‘The Problem’ above), but both are guilty of exporting their own ‘gene-centric’ conception beyond the boundary of its application.
This needs some unpacking.
By shortening the recombining units involved in the basic transaction, we have dropped down to a new, subgenome level of selection. In a perpetual diploid, the entire paired genome is subject to repeat testing en bloc. But by continuing to reduce to haploids in sex, each chromosome is selected independently, for combinations that work together along its length but in competition with other such chromosomes segregating in the diploid population. Even though we have not yet introduced intrachromosomal segmentation by crossover, we have all the conditions required for ‘selfish genes’.
Dawkins’s catchphrase is frequently misunderstood, in both its parts, though he takes great pains to explain himself. ‘Selfish’ sets up the paradox of his most famous book: how to explain altruism in a competitive, Darwinian framework. His ‘Gene’, meanwhile, is not the molecular biologist’s or the geneticist’s, but a shorthand for a recombinant unit – a stretch of genome bounded by the extent to which it independently recombines. Maynard Smith’s ‘gene A‘ is not a recombinant unit; such things do not exist in asexuals.
Selfish genes integrate into the genomes of the future diploid population; that integration depends entirely upon recombination, and hence upon sex. Along their length, shorter stretches are selected to co-operate, because their futures are linked. While in the diploid state, all genes again have a common interest in co-operation. However, on separation, selfish genes are supposed to oppose that separation. But this is to misunderstand the causal mechanism underlying the metaphor. How can they do so while still remaining selfish genes? While it can be helpful to think in terms of a gene’s ‘wishes’, the metaphor breaks down the moment their means of population integration is removed, and becomes misleading. Paradoxically, a selfish gene cannot ‘wish’ to stop being a selfish gene!
Further, genetic loci themselves are not omnipotent. Most genes supply proteins or RNA tasked with performing some biochemical function or another. This is not readily modified to do something completely different – to ‘plug up the works’ of meiosis in some way. It is simply not mechanically possible for most genes to directly influence their transmission. They must do so through their effects on fitness, in whatever genetic system they find themselves. Indeed genes – the majority – which are at high frequency in the population gain no copies by abandoning sex, since they will generally be homozygous most of the time in any case.
Therefore only a handful of genes have even the notional capacity to effect or benefit from the switch to asexuality. Such genes would leave the sexual population, taking everything else with them – they become de facto a separate species, imprisoning a genome which can compete as a lumpen diploid whole, but whose parts can no longer integrate ‘selfishly’.
This is not to say that mutation to asexuality cannot occur, but it is an error to see this as a genetic competition between genome subunits, when it completely removes the sense in which such subunits have ‘interests’. It is an ecological competition between species.
The only lever available to recombining genetic stretches is to distort their transmission in meiosis, by tinkering with its machinery or attacking their homologue, rather than abandoning meiosis completely. Even here, mechanisms are dissipative. While such drive may distort transmission from an equitable 50/50 for a period, when a distorter becomes common it starts to encounter copies of itself, and either transmission returns to 50/50, or the distorter suffers from attacks upon itself in homozygotes.
Advocates of selfish gene viewpoints sometimes imbue them with a reach that exceeds their grasp. Far from being ‘dragged kicking and screaming’ into meiosis, they file obediently in, as they have always done.
Genetic algorithms
These are computational search heuristics inspired by biological population processes. Digital chromosomes representing varying solutions to a problem are copied and varied further, and the ‘fitness’ of the population members evaluated to determine which persist into the next round. Adding recombination to such programs can have a dramatic effect on search times, and aid escape from local fitness maxima. Recombination is gentler, less ‘speculative’ than mutation, since both parts have already survived in the population.
Even though evolution is not a search as such, the effects of recombination on rate and connectedness in GAs must surely have an analogue in the evolutionary behaviour of the natural populations from which they take their inspiration.
______________________
4) Speciation
So far we have dealt with a single species – the only ‘true species’ on earth under the Biological Species Concept, everything else being an asexual prokaryote or eukaryote. Over time, any persistent reduction in free gene flow around such a population would be expected to lead to divergence between the subpopulations so formed – ultimately to the point at which the subpopulations would be incompatible, due to biological isolating mechanisms. At such a point, the sexual clade would have broadened, and the supposed broad-scale threat of asexuality, to extinguish all parent species, reduced. If there is a probability p that any one species will be eliminated by its secondary asexuals in a given time, with two species the probability they both go is p x p, with 3 species p x p x p and so on. The longer sex survives, the more resilient it becomes, simply by putting its eggs in multiple baskets.
Furthermore, once the growing clade had acquired a degree of ecological divergence, new asexuals would have more than just the resident competing sexual parent species to deal with. The sexual clade may begin to throw up predators, prey, parasites and interspecific competitors, all of which possess greater variation and evolutionary fleet-footedness compared to each new clonal variant. This is termed the Red Queen effect – it takes all your running just to stand still! – and further serves to cement the sexual clade’s position. Secondary asexuals surely arise from time to time, but most either fail to supplant the resident, or succumb to extinction by the several forces ranged against them after replacing the parent species.
As to the original haploid populations from which sex sprang, they too suffer from attrition by the sexual clade. The dynamics provided by sex are not only manifest in the diploid phase. The sexual clade can tune haploid genomes at chromosome level (or lower, given crossover). It can generate beneficial combinations of genes, and possess greater variation even within the haploid phase of its cycle than is available to perpetually asexual haploids. All of these, when added to advantages in the diploid phase, would tend to see gradual extinction of the ancestral asexual haploids.
5) Crossover
Some of the effects of recombination are available passively through independent segregation of chromosomes, discussed above. A distinctive feature of modern sex, however, is further segmentation of chromosomes through crossover. This involves reciprocal swapping of segments of homologous chromosomes during separation in meiosis. A nick is made in one chromosome, the homologue is recruited to provide a bridging patch, just as in repair of accidental breakage, and the resulting interlinked structure is resolved to yield separate chromosomes. There are four different ways to resolve, two of which yield recombinant chromosomes and two return the chromosomes unchanged (but for the patch, which results in gene conversion).
In the ancient repair system from which this derives, the random swap/no-swap result made no difference – the input chromosomes were the identical sisters freshly duplicated during the S phase of mitosis. But with nonidentical homologues from separate parents, and independent futures for the outputs, the consequences are profound.
Because crossover sites are not fixed, it has the effect over multiple generations of exposing yet shorter genetic stretches to independent selection – fully unmasking Dawkins’s ‘selfish genes’, each locus uncoupled from its neighbours and tested independently, over evolutionary time. As with segregational recombination, we get
- Increase of beneficial alleles and decrease of detrimental with reduced interference between loci.
- More rapid creation of novel combinations
- An increase in variation, promoting both local adaptation and evolutionary resilience
These are arguably the most significant consequences of sex, which indeed cause many authors to regard it as the whole ‘point’ of the enterprise; the reason it exists. However, on the argument presented here, this need not be the case. After all, if the gene conversion and segregational recombination above were purely incidental, might it not be the case here too? 50% of crossover sites resolve to a recombinant product, but genes are blind to this. No gene is fundamentally bothered whether it remains linked to the same or to different chromosome-mates, provided the result works.
Population effects are important for lineages, but we do need to get active recombination from low to high frequency before its population consequences can be manifest. Felsenstein (yes, that Felsenstein) considers the case in which that may be mediated initially by drift, subsequently cemented by the resilience of such populations to environmental change. My own preference is to appeal to cellular mechanics. Crossovers assist in the equal tensioning of homologues as they are hauled apart during meiosis. Without them, there is a tendency to damaging asymmetry – one haploid output may lack an entire chromosome, the other having two copies giving potentially damaging trisomies on subsequent fusion. This gives a sufficient reason for crossover to become common in a sexual population without appeal to circumstantial issues such as environmental fluctuation or the extent of negative epistasis in the population. While recombination has significant population effects, which play a substantial role in the success of the clade, those consequences may be arguably a side-effect of its cytological role – an example of what Stephen Jay Gould termed a ‘spandrel‘.
______________________
Intermission 3 – macroevolutionary trends
As a result of these various dynamics, we might expect gradual elimination of the ancestral asexual haploid lineages, possibly complete even prior to our Last Eukaryote Common Ancestor. Certainly, there are no known asexual haploid lineages today.
Since sex has its roots in the prehistory of the entire eukaryote clade, it probably played a significant role in eukaryogenesis itself, that mysterious and probably extended sequence of amendments separating eukaryotes from prokaryotes. We see that as a singularity because we have no surviving intermediates. But sex – serial diploidy – may be at least as important as endosymbiosis, the origin of mitochondria, in the overall sequence. In mitochondria, we are familiar with two genomes in a cell, but in reality, in the diploid phase, there are three.
Subsequent to LECA, the constraints on invasion by asexual diploids and the greater capacity of sexual lineages for anagenesis and cladogenesis would lead us to expect to find a eukaryote clade consisting mostly of sexual forms, with few if any purely asexual haploids, and comparatively few secondarily asexual diploid species. Which is handy, because that’s exactly what we find! All of this was achieved without going anywhere near any ‘twofold cost’. Isogamy can get us a long way.
______________________
6) Multicellularity
Multicellular organisms, featuring multiple tissues in one genetic ‘individual’, have arisen independently a dozen or more times in eukaryotes, but never in prokaryotes. Because the eukaryote clade is built upon sexual roots, there is a close relationship between multicellularity and sex. After all, taking the ‘haploids’-eye-view’, what is a multicellular body other than a vehicle to protect and amplify both genomes, before returning haploid copies in profusion?
Of course there is a little more to it than that.
A multicellular eukaryote forms from either or both of the haploid or the diploid state, according to life cycle, by repeated mitosis of the genome of one or a few cells. The multicellular mode gives advantages of size and of ‘division of labour’ – by differential gene expression, different cell types can perform different functions despite possessing identical genomes. Among those specialised functions is reproduction itself. The cells of a body, or the genes within them, forego their direct reproduction in favour of the reproduction of identical gene copies in the specialised reproductive tissues. Freed of the need to reproduce directly, other tissue cells can concentrate on their own function.
While haploid individuals do occur – for example male social bees – the predominant mode is to form a multicellular soma from the diploid. Even fungal fruiting bodies, which avoid true diploidy until spore formation, have cells that contain both haploid nuclei: dikaryons. Enforcing a reproductive dead-end on diploid cells (or fungal dikaryons) may be easier to orchestrate, since reproduction is performed via production of a non-diploid cell type, a specialism not easily accessed by cells specialised for other functions. (Male bees may be exceptional because they are functionally the sperm of a ‘superorganism’: the hive).
Thus , as a general pattern, the co-operation of all cells in transmission of their shared genome appears to be secured by the specialised exit; opportunities for rogue cell lines to ‘go it alone’ are reduced. Conversely, a hypothetical asexual lineage exploring the first steps towards multicellularity has no equivalent mechanism ensuring intercellular co-operation and specialisation of the reproductive function. Multicellular bodies, I would contend, are an invention of sexual lineages.
7) Gender
The key distinction of ‘male’ vs ‘female’ is relative gamete size, rather than the organs, tissues and bodies with which we are more familiar. Females have the larger gametes, males the smaller. Note that, genetically, there is still little or no distinction between the haploid gametes; it is purely a matter of cellular packaging, with female gametes getting the lion’s share of cytoplasm. For this reason, it is unlikely that gender so defined can arise in unicellular organisms, by asymmetric division; the smaller would suffer disproportionate losses, and any genetic conflict would appear to centre on equal division. By generating male gametes in multicellular organisms from the end of a series of diploid mitoses, however, we get massive amplification of the genome, compensating losses by sheer weight of number. Female gametes, meanwhile, can be nourished, and furnished with additional cytoplasm to kick-start the next generation. Male gametes, being smaller, do most of the dispersing. Being cheaper to produce, they can be generated in greater numbers, but ultimately offspring numbers are limited by females.
______________________
Intermission 4
Cost of Males
Leaving aside, for want of space, the reasons why gamete asymmetry is more stable than isogamy in multicellular organisms, we are finally in a position to examine the ‘twofold cost of males’. Note that this is a long way down the evolutionary trajectory. We have managed to get almost every feature of a modern sexual system, without a whisper of a roadblock from that twofold cost which is supposed to render sex itself a mystery. Even now, it is only an issue for dioecious organisms – those with separate male and female individuals. So the ‘mystery of sex’ resolves to a mere ‘mystery of dioecy’.
Imagine first a multicellular species with slight asymmetry: ‘male’ gametes are fractionally smaller than ‘female’. Fertilisation takes place externally. An asexual offshoot of the female version could, by abandoning fusion, produce asexual diploid offspring directly. But what would such a lineage gain? As in the primitive scenario, all asexual mutations occur against a background of a resident sexual, having variation and faster evolution at its disposal. The asexual is not helped in this conflict by the supposed ‘twofold cost’. We can incrementally increase the cytoplasmic asymmetry, but there is no clear point at which the need for an individual twofold benefit to sex arises. Nor does it arise incrementally, in proportion to the increase in gamete asymmetry. The relative benefit of sex is ‘smeared out’ among the population members, residing in variation already generated, in future capacity for tuning, and in the minimisation of gene conversion-induced homozygous effects.
Finally, we can add resource asymmetry in the embryo to the picture. Females often provide far more to the next generation than males, much more than just a bit of cytoplasm. While males continue to offer nothing more than haploid genome copies, as sperm, spores or pollen, the developing embryo may remain part of the female, as a baby, a maturing fertilised egg or a ripening seed. Half of those babies, eggs or seeds are male. The opportunity finally arises to increase ‘twofold’, by producing only female offspring. This would result in twice as many grandchildren, and exponential increase, compared to the sexual. However, again asexuality happens, when it happens, against a varied sexual background. It is still by no means certain that asexuality should be expected to win this contest a sufficient amount of time to eliminate all parent populations.
The gene’s perspective, again.
Gender is about resource asymmetry, not genetic asymmetry. Most genes reside on autosomes, and as such spend half of their existence in each body, investing equally in two complementary strategies. While in a male, they get inserted into large numbers of mobile gametes, widely dispersed. While in a female, they get inserted into fewer, less mobile but larger and better-provisioned gametes.
Despite several added complexities, from the perspective of any given gene the situation has still not changed from the single-celled state sketched above. In that sketch, individual genes feel no ‘force’ compelling them to remain paired indefinitely – to become asexual diploids. They come together for a period, then part. Even with full recombination, as long as the upstream and downstream companions of any gene constitute a viable haploid, it doesn’t matter whether those companions came from the same or different input haploids. In adding crossover, multicellularity and gender, we have changed nothing in terms of the dynamic between the haploid and diploid states. A given gene in the simple cartoon cycle above will appear in 50% of haploids, deterministically. In the more complex state we are now considering, a gene has exactly the same odds, now offered stochastically.
Haploids go in two by two, haploids come out two by two. Their roles as units are somewhat obscured by their loss of individual integrity as a consequence of the amplification and ‘randomisation’ of the input genomes. With independent segregation and crossover, output haploids are scrambled versions of the inputs – every one of the sometimes billions of outputs is unique; a sexual snowflake. But there is no gene in the haploid which is fundamentally motivated to ‘object’ to this scrambling. Throughout, it remains a genetically symmetrical transaction of pairing and parting.
______________________
Population Genetics.
The ‘twofold cost’, along with other proposed costs of sex, such as recombination and segregation loads, is an argument derived initially from population genetics rather than observation.
Population genetics attempts to model populations in terms of mathematical or computational abstractions – necessarily, simplifications. The population in a simple equation is considered to be panmictic – all individuals are equidistant from all others, and equally likely to mate. Note here that sex is built into the very fabric of the assumptions. The panmictic population is stirred by, among other things, sex itself, through mate search and gamete dispersal. Such populations are also bounded by sex. If an obligate sexual, the range is determined in part by the need for a partner. A male can wander freely, but his genes cannot be passed on if he wanders too far from the available females. The same applies to an unfertilised female.
Now, population genetic models can be applied to asexuals. But there’s a bit of a difficulty there, because they lack both the stirring and bounding effects of sex. We artificially assume they are stirred and bounded, even though a significant cause of these is absent: sex itself. This is OK(ish) until we try and use the same model to compare sex and no-sex. Maynard Smith makes the same error in his textual argument above.
Consider what we’re doing when we use an equation to model a population. In essence, we are putting the population into a massive corral and giving it a good old stir. Individuals can leave the corral – it’s more a line in the sand than a fence – but they can only breed within it. Now, we add a few hypothetical asexuals. But implicitly we keep stirring at the same rate – despite the asexuals not being subject to those ‘stirring’ vectors related to sex. We also keep the same ‘breeding boundary’ – despite the asexuals not being constrained by it.
In a real population in which you bodily moved every new asexual individual to a different location, and kept them corralled, and eliminated all variation in the parent sexuals … then yes, the ‘cost of males’ might be an issue! The simplification is equivalent to metastasis in cancer. If cancer cells were always spread evenly around the body, cancer would kill in far greater numbers.
Because asexual individuals do not disperse in the same way as sexual genes, it is misleading to count their numbers as if they did. If an asexual colony arose within the ‘corral’, partially displacing the resident sexual locally, many of its offspring would be competing not with sexuals, but with other asexuals, reducing the impact of asexuality from that in a maximally-mixed model.
Asexuals are also free to leave the ‘corral’ entirely – here they find no competition with sexuals. Yet the model metaphorically rounds up such individuals and shoves them back inside, efficiently and artificially mixing each such individual to a different location within.
Using a simple model substantially over-counts asexuals.
How easy is it?
A fundamental assumption of ‘cost’ models is that the mutation to asexuality is freely available. This may not be the case. Clearly, to be costly, something must exist that can cash in on the saving. In a land without predators, it costs nothing for a zebra to browse leisurely and alone. Likewise, sex is only costly in the face of an existent asexual.
Firstly, mutation rate is proportional to population size. Small populations are most at risk from asexual invasion, as they lack variation, but equally they are less likely to suffer the mutation(s) in the first place.
Secondly, there is not a uniform susceptibility to the mutation across the tree of life. There are, for example, no known asexual mammals. There may be several reasons for this, but difficulty of mutation may certainly be one. Sperm import imprinted genes which have a profound effect on embryogenesis, while female meiosis is geared towards sperm reception. For a female mammal, it may not be a simple matter of ‘turning off meiosis’. She cannot perform autogamy either, fusing her own gametes, since 3 of the 4 outputs of meiosis are shrivelled ‘polar bodies’. Female mammals, with internal fertilisation, embryo retention and post-partum nurturing might seem to have the most to gain from asexuality, but they may also, as a group, find that amendment the hardest to achieve. This, rather than cryptic twofold benefit, may account for its absence.
I am only aware of one asexual bird, the turkey. And those offspring are all male, and so can’t form a lineage.
Asexuality becomes commoner as we go through the reptiles, and on to fish. However, even there, many of the examples arise from a process known as hybridogenesis rather than mutation. A significant competitive difficulty facing hybridogenesis is the replacement of both parental species, if they are ecologically distinct. Therefore, such mechanisms don’t pose a great threat to sex on the grand scale.
______________________
To summarise, sex is woven into the fabric of the whole of eukaryote biology. It is an error to view asexuality as if it were a simple adaptation for diploids; much of the ongoing puzzlement derives from adherence to that standpoint. Contingency, side-effect, central stability and exaptation play at least as great a part as adaptation in the prevalence of sex, while the expectation that a twofold benefit is still required is an artefact of the diploid stance in oversimplified models. Sex and diploidy are fundamentally an adaptation for haploids, with far-reaching consequences.
It seems reproduction isn’t exactly what we have always been told. I learned just recently that if a woman has more than one partner just prior to conception (within days I believe) that the child can get genetic material from both men, even though only one sperm pregnates ! Now when did school teach that?
phoodoo,
Sperm mediated gene transfer.
Allan Miller,
I am not sure I understand this link, I don’t think that is exactly what I am talking about.
https://time.com/3461485/how-previous-sexual-partners-affect-offspring/
This article was written in 2014, talking about the effect in flies. In the article it mentions they are unsure if this is possible in humans. But from what I understand, we now know, that indeed, other sexual partners do affect the future offspring.
phoodoo,
It’s a possible mechanism. SMGT exploits the tendency of exogenous DNA to stick to sperm, and hence be injected into the egg along with the sperm’s contents. What I’d surmise happens is that, with multiple partners, rupture of some sperm from one partner may lead to some of its DNA sticking to the sperm of the other.
phoodoo,
Although, reading your link, they seem to be talking about epigenetic effects, so it may be inapplicable.
Allan Miller,
Yes. I will have to find the newer article I read. It’s not about previous sperm hitching a ride onto the impregnating sperm.
To do what? To confuse the public?
Give me something to contribute to other than: ‘it must’ve happened that way or else evolution is toast’…
Yeah… the lack of evidence for your speculative ideas…
So you made it up? Impressive!
Really?!
Are you questioning the experimental evidence for quantum jitters in mutations?
If yes, based on what evidence? Your imaginary nurturing and bathing processes in the cell?
Maybe they are quantum? 🤣
You are blaming Allan for your strange impotence to phrase an intelligible objection?
And another one. You have posted half a dozen comments with the word “speculation” in, but consistently fail to specify what exactly it is you object to.
This is all very amusing, but you don’t have the faintest understanding of what the OP is about, except that it involves evolution, do you?
And you and Alan have posted what in response???
Nevermind!
The usual narratives that make you feel like you’ve accomplished something nothing to do with real science…
Pity…
This one bothered me, so I did look up some studies and found an interesting paper. It supports your view by showing that sexual yeast cultures have more rapid local adaptation in an experimental microcosm experiment in the presence of gene flow. Interesting is that they find this depends on the genetic architecture of local adaptation. If local adaptation relies on antagonistic pleiotropic alleles with opposite effects in different environments, then sexuals suffer from high migration rates. If however, as was probably the case in the experiment, local adaptation proceeds by purging locally maladaptive alleles, sexuals have an advantage, because they can rapidly break down LD beween adaptive and maladaptive alleles.
We are comparing the two reproductive modes. Mate limitation is definitely a cost compared to asexuals, for example at low population density.
But sometimes they won’t. For example, Here is a paper on evening primroses, demonstrating that functionally asexual lineages indeed have slightly higher extinction rates, but this is more than compensated by much higher diversification rates. The latter makes a lot of sense, because asexuals lack the homogenizing effect of sexual recombination and segregation. That’s just one example of course. But it shows that life doesn’t always behave like we expect it should. I really need to see increased asexual extinction rates demonstrated, before I accept them as fact. That’s how I roll.
If asexuals live in places where sexuals can’t, it demonstrates the failure of sexuals to compete in those places. I’d like to see some data on ranges of sexual and asexual sister species by the way. I am not convinced they do not have overlapping ranges.
Be careful what you wish for. Finished your OP though 🎉.
Requests to specify what you object to … which you ignored.
Is it possible that sex is just the result of drift?
No strong advantage or disadvantage?
Just happened and survived?
I’d be inclined to think that the latter is the norm. If ‘local’ alleles or positive epistases have been concentrated by selection in sexuals in the first place, that same process is ready and able to keep going. Migrant alleles are indistinguishable from the alleles that were substituted during initial adaptation.
What we are looking for is an overall assessment of how likely it is that asexuals can replace the resident sexual through evading ‘breakup’. The complete dynamic needs consideration. There will be a rate of reversion to asexuality. Being smaller than the entire population, any given deme is less likely to experience it than the population as a whole. Different alleles will have different geographic ranges over which their ‘local adaptation effect’ is positive – a deme for one allele could be a ‘subdeme’ of another. Some alleles will be adaptive wherever they occur – the set of all demes. Abandoning sex cuts one off from all of this network – as well as reducing the efficiency of any selective sweeps locally in train, due to interference.
Models that assume unlimited production of asexuals wherever required, neat circumscription of a particular set of adaptive alleles to one range while ignoring all others, and what seem like quite strong selection coefficients for the combinations (if that’s all it takes to eliminate sexuality!), will greatly overestimate this ‘cost of sex’.
Not if there’s nothing else about to reap it. On the one hand you’ve invoked low transition rates as a possible explanation for taxonomic distributions, but now you make an assumption that has asexuality available continuously. We humans stare down at the process and say ‘look, you could just mutate and do x’. But until something starts doing x, there is no ‘cost of not doing x’. “Just you wait till something starts doing x, then you’ll be sorry”. 😃
If we were to compare the cost of different residence times at a watering hole for zebras, we assume the existence both of something that causes differential reward, and of the variants themselves. If we remove the predators, we remove the discriminator. If we remove the polymorphism, we remove the differential. Cost of sex thinking in general imports both ‘predators’ and ‘polymorphism’ rather indiscriminately.
But you’re happy to accept models for the alternatives? I don’t think, even if I demonstrated it once, that would be conclusive. You could hang on to your skepticism through quite a few such empirical tests!
I was talking of that particular model. It just seems an odd choice of ‘neutral’ model to me – “look at how well asexuals do in these marginal habitats that lead to their elevated extinction …!”. Why don’t they live somewhere better? They can’t, something’s already in residence.
I think there’s an element of truth in that, though we do need an explanation as to how it got going and why almost everything still does it (or their recent ancestors did). Drift won’t do it, but contingency and incidental effects do have a strong part to play, I think. The transaction is inherently stable, and highly resistant to invasion by secondary asexuals (those of sexual ancestry)***, rather than being generally ‘advantageous’.
*** Not everyone agrees, of course!
“Drift won’t do it” sounds like an Edge argument.
On the other hand, trying to find the origin of sex sounds as difficult as finding the origin of life.
It needs more than neutrality to get sex going in a population of asexual haploids. There needs to be some advantage for something.
More like as difficult as finding the origin of eukaryotes. Anything that is shared by all eukaryotes must have happened prior to their last common ancestor. That includes sex, but also mitosis, mitochondria, cytoskeleton, loss of cell wall, nucleation … but we only see a singularity, because any hypothetical intermediates are extinct except for that one lineage. We don’t know the conditions of the specific species at the time – and it was just one species in a tiny corner of the planet – so can only take a guess based on reasoned argument.
My hypothetical scenario emphasises the importance of diploid formation in a population of haploids. That could be advantageous (and hence not down to drift) because of size increase and/or hybrid vigour, for example. The advantage need not be huge, just appreciable, to compete with the ancestral population of haploids. At some point prior to the present, and possibly prior even to our common ancestor, all asexual haploids went extinct, IMO because the sexual clade diversified, gained variation, and was able to ‘tune’ genomes at a lower level, and hence more efficiently.
Since then, the clade has diversified still further, the original reasons for cyclic diploidy being largely superseded by others, such as secondary consequences that work at population level, and the production of large multicellular somas like ourselves.
Corneel,
You’re right to challenge me to put my money where my mouth is – to find some direct evidence that competitive exclusion differentially favours the sexual resident – but I can easily make the reverse demand. Why is sex deemed mysterious? Because models. Asexuality typically wins, when you flatten everything out. So does that actually happen in life? No, because ‘cryptic benefit’…
‘Cost of sex’ thinking is largely based on population-genetic considerations and assumptions, rather than empirical observation. So what should persuade me to drop my opposition to that stance – my criticism of the models used and the conclusions drawn, that on the basis of these models, sex must offer a cryptic benefit in order to survive in the real world?
I have repeatedly toyed with the idea of ‘a better model’. I think geography needs including, and population density, rate of mutation to asexuality, some kind of heterogeneity of environment across the range, a decent set of alleles segregating in the population, cladogenesis, a sexual ecosystem, drop the assumption of panmixis but add the vector of male dispersal. Sex, I am convinced, will do better and better as the model becomes more and more ‘life-like’ – and computationally prohibitive! The more ‘reality’ I put in, the more I’d be accused of gaming the setup – building a model that favours sex!
Yeah, that’s why I write … 🙄
You surely don’t think I imagine the female gamete as equivalent to a nursing mother at bath-time, do you? 🤣
Female gametes provide the bulk of the cytoplasm in the zygote. It’s how the male/female distinction is defined: relative gamete size.
Random genetic drift is, well, random. How could it?
Is there an alternative to the view that sex has to offer some advantage that asexuality doesn’t?
Drift is an important element, though. Just not the whole thing. It can allow alleles with little individual effect to become common enough such that population effects become significant.
I’d agree, but tightly linked loci in repulsion phase may be indistinguishable from antagonistic pleiotropic loci. Again, this is likely to come down to specific details about recombination rate and genetic architecture of local adaptation.
And I am willing to bet there are several taxa out there where these assumptions are reasonable. Problem is, we are confusing origin of sex with maintenance of sex again. My personal view is that in general, sex is maintained by it being the ancestral condition, and most lineages being locked into sexual reproduction mode.
Science is conservative. Deal with it, you sexual chauvinist!
As I said, I am not sure whether asexual and sexual ranges truly are non-overlapping. If there are ranges where they live happily together in most cases, this argument will look different. If I have time, I’ll have another shot at finding those data (didn’t succeed last time).
There are lots of empirical observations, but they are typically non-conclusive because we can only test in highly derived eukaryotic lineages. I accept your argument that for the origin of sex, we should disregard complications of anisogamy and dioecy. For the maintenance of sex in extant lineages, I maintain this will be quite relevant though. Sex simply opens up a playing field for all kind of bizarre conflicts, which are bound to result in costs at the population level.
Welcome to the club, petrushka! 😁
This is definitely something I would write…
With natural selection getting the beating from the leading evolutionary biologists, like Michael Lynch, for the claims of an all-powerful mechanism:
“…evolutionary biology becomes little more than a (sometimes endless) exercise in dreaming up the supposed agents of selection molding one’s favorite aspect of phenotypic diversity. …
… we now know that this unwavering belief in the limitless power of natural selection is untenable…”
https://www.sciencedirect.com/science/article/abs/pii/S002228362030156X?via=ihub%22%20%5Cl%20%22!
…what’s left?
Random mutations that are not really that random and require quantum jitters; quantum information-the rearrangement of quantum states to happen? 😉
Darwinists had better begin learning quantum biology before they embarrass themselves in public, like Swamidass has recently…
Yeah, you confuse more… You can’t even get Darwinists on your side; i.e. Harshman…
No? That’s fantastic! But you’d better have a mechanism for this IMAGINARY FRIEND, before you publish it…
No way! And this is what you call nourishing and bathing? 🤣
So which of us is right, him or me? Whichever one disagrees with the other? That’s both of us!
What in hell are you on about?
Not sure I used the word ‘bathing’, but it does sit around the egg’s genome, and the male’s, post-fusion, so I wouldn’t think it outrageous to do so. Cytoplasm from the egg supplies the bulk of the nutriment of the zygote. That’s ‘nourishing’. I don’t see what objection you are raising over me saying that. What do you think cytoplasm does, if it doesn’t surround genomes and supply their initial nutrition? Are you thinking of ectoplasm? 🤣
For sure, but the ‘mystery of sex’ idea is framed in far more general terms.
I’m not keen on that dichotomy myself. When do you think ‘origin’ stops and ‘maintenance’ begins? It’s as if people think there should have been just enough sex to make diploids, then go back to asexuality again, without something ‘maintaining’ it. Would that extinction of sex happen before or after dioecious multicellularity evolved?
That’s part of it, but can’t be the whole. Extrinsic factors, such as the inevitable tenure of sexuals, and the rate of production of asexual mutations, must be important, but you can’t sweep the intrinsic ones under the carpet: competitive difficulties, the reasonable possibility of genomic deterioration, interference with selection and evolutionary rate.
Diplocentric splitter!
Survivorship bias, though …
Yeah, now! But it started on paper.
It’s also inevitable that model organisms have overcome some barriers to give detectable numbers of asexuals. This doesn’t help with getting a handle on overall tempo.
It’s true, but I don’t think these conflicts ‘select for’ the abandonment of sex. Meiotic drive, for example, selects for modifiers within the parting haploid genomes, and dissipates when the driving allele becomes common. But a lot of people see that as a cost of meiosis. It only is, even notionally, while the driver is rare, so we’re assuming that’s enough for asexual replacement, which is doubtful. And regardless, an asexual could evade drive, but access a whole bunch of other problems instead. No-one talks about the costs of no-sex! Cos you can’t stick a number on ’em! ‘Twofold cost’ has become a slogan.
I think in general the problem seems to me to revolve around people conceptualizing sex as an allelic ‘switch’. It is much more than that: it changes the very nature of alleles. Things don’t just carry on as they were, except for the fact they don’t do sex anymore. They’re a different species, with a very different mode of genetic assortment.
Is there a third option?
I was asking you…just in case you haven’t noticed.😉
This pretty much sums it up…🤗
This is the mechanism? Cytoplasm supplies nutrition?
That’s fantastic! Why didn’t you say so in the first place? 😅
But, I doubt John Harshman is going to buy this as a mechanism leading to the evolution of sex… John may be reading and studing the bible, but i don’t think he is a man of that kind of faith… Even he knows where to draw the line…😉
How about this? Thermo-vents?
Or a bolt of lightning? Take your pick…🤗
No
Because it has no relevance ‘in the first place’. It relates to gender, which is item 7 in my proposed sequence. Even then, it’s definitional, not theoretical.
Well, that’s not my mechanism, but – so? Is he the arbiter, now? There is an entrenched position in biology in regard to analysis that I am arguing against. John, and to some extent, Corneel, represent that position. So naturally they are going to defend it. That doesn’t make me wrong (necessarily), just unpersuasive.
But at least they won’t embarrass themselves with the comprehension problems you are displaying, obsessing over a couple of concepts appearing very late in the argument as if they were the argument.
And what’s the meaning of this pure gibberish? As the saying goes, you should have remained silent…
Physician, heal thyself! 🙂
I’ll give you that if you can translate this from nonsense into logic:
Two parents contribute half a genome each to offspring.
J-Mac,
Welcome to the club, petrushka! 😁
This is definitely something I would write…
In a parallel universe, perhaps, but you would not mean what I meant.
Hardly my fault you don’t, and won’t, understand genetics, is it?
That’s not quite the whole story though. Without sex, all the chromosomes in a cell stick together in the same lineage, encapsulated and hence linked through the generations by the cell membrane. With sex, they travel independently down the generations due to independent segregation, and therefore there is a new level of selection. Genes are linked covalently on any one chromosome, but sex can sort those chromosomes at a subgenome level, into separate lineages.
With sex and crossover, the level of selection becomes even smaller. Chromosome fragments – alleles – travel independently, according to the extent that recombination occurs between them. The covalent linkage is broken, then reformed, in crossover, and half the time the result is a swap. Sex is now sorting at a subchromosome level.
Am I correct to think that a sexually reproducing population has effectively more variation, because of recombination?
Generally, yes, though it depends on the extent of inbreeding and bottlenecking.
But for example if you have 2 chromosomes in the haploid, a population which consisted of A/B or a/b asexual genotypes – 2 varieties – can turn that 2 into 4 simply by fusing then separating. The haploid population would have A/B, a/b, A/b and a/B individuals, purely by accident.
Variation becomes greater still with the further subdivision provided by crossover – though I don’t think that’s the prime reason for crossover.
You also get less ‘hitchhiking’ with selection when recombining units are smaller, which can be a source of variation loss. Selection eliminates variation (as does drift); the bigger the units, the more variation lost.
Nonlin.org,
I had no trouble understanding Allan’s point. Your inability to understand simple concepts in genetics may be rectifiable, but only if you are willing to learn.
The auspices are not good.
So? So you are taking it back? So what?
We have Allans to pick on…🤣
I really like when people like you use this excuse, when cornered…It makes them feel alive…again…or something… 😅
Drift is not the result of differential reproductive success, but individuals must survive. That necessity is a kind of selection.
Once you have passed through a door, new avenues exist. The drunkard’s walk moves you inexorably further from your origin.
My point is that change in populations is not necessarily improvement or progress. It is change. Sometimes you can observe adaptation, but evolution will change populations without differential success.
Sex matters in perspective 😎
True enough, but each time a change arises that affects success differentially, it tends to be fixed or eliminated more strongly than those that don’t. So there is a bias in favour of those fixations. There is no good measure of ‘improvement’ or ‘progress’ itself. If a 10,000-strong population fixes an adaptation, and ends up still 10,000 strong, it has not obviously gone anywhere ‘better’- although it might have, if that population has become a shade less likely to go extinct due to external factors. That’s really the measure of success at all levels, from clade to gene: sticking around. Some organisms stick around by splitting every 20 minutes or so. Others produce millions or billions of gametes just to generate a couple of offspring over 70 years or so. Neither strategy is intrinsically ‘better’; they both work.
It will indeed.
I agree too but in small populations, random variation will produce new alleles and drift will fix some and eliminate others randomly. Change, yes, but without adaptations, what’s the trend? Extinction?
More likely extinction, though no guarantees. New World monkeys, for example, may have evolved from a single pregnant female, discussed by Dawkins in The Ancestor’s Tale.
It ties up to the discussion with Corneel above. Partly informed by such issues as inbreeding depression and variation loss in endangered species, I feel that a clonal asexual will struggle against a varied resident for that reason alone. Gene conversion in recombinational repair provides a close analogue of inbreeding, by tending to increase homozygosity the longer a pairing lasts.
Drift is a useful ‘force’ though. When an allele goes from single copy to fixation, spread may be all-drift initially, but with increase, density- or frequency- related effects can speed it on its way selectively.
Except one produces more of the same while the other produces self-conscious, creative individuals with unique biographies and with a measure of being able to tell right from wrong, good from evil. Although the ability to make these distinctions does not imply taking the appropriate course of action.
Of course for those who believe that it’s all down to natural selection acting on random changes neither of these strategies has more value than the other. When it comes right down to it, life is one big grey amorphous meaningless mass spreading over the lifeless matter of the earth like an invading cancer.
Well duh. And what is the connection with “descent with modification”?!? I read that nonsense and it doesn’t translate into what you’re interpreting in any language. Obviously we’re a mix of both parents and NOT a modified version of either. So your “interpretation” is divorced from “descent with modification”. This has been discussed… at nauseam.
And where is the genetics in “descent with modification”? See above including “at nauseam”.
See above. I dare you to link genetics with “evolution” in a logical, intelligible sentence.
Sure, for anyone who believes the measure of success is CharlieM …
Yeah, that’s why I kick tramps and babies. ‘Cos of biology.
Evolution is descent with modification; genetics deals with both the descent and the modification parts. I really don’t know why you have such trouble with these concepts, but the way you climb atop your molehill and declare to the world “Evolution Has Nothing To Do With Genetics” is, to any informed reader, utterly hee-la-rious.