
As shown repeatedly, “Nothing in Evolution Makes Sense. Period.” Not natural selection, gradualism, human evolution, UCD, tree of life, etc. And just to confirm, let’s look at another one of the nonsensical concepts of “evolution”.

- ‘Divergence of character’ (character displacement or sympatric speciation) postulates: “during the incessant struggle of all species to increase in numbers, the more diversified these descendants become, the better will be their chance of succeeding in the battle of life. Thus the small differences distinguishing varieties of the same species, will steadily tend to increase till they come to equal the greater differences between species of the same genus, or even of distinct genera” (Darwin 1859). Sympatric speciation is hypothesized as “the evolution of a new species from a surviving ancestral species while both continue to inhabit the same geographic region”.
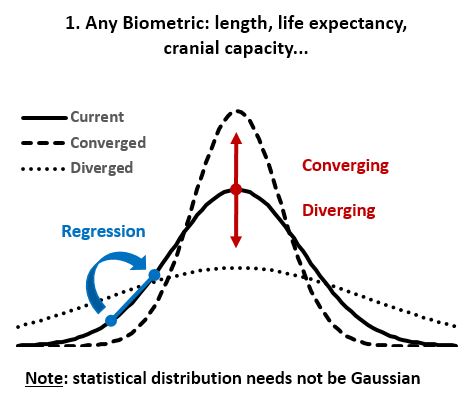
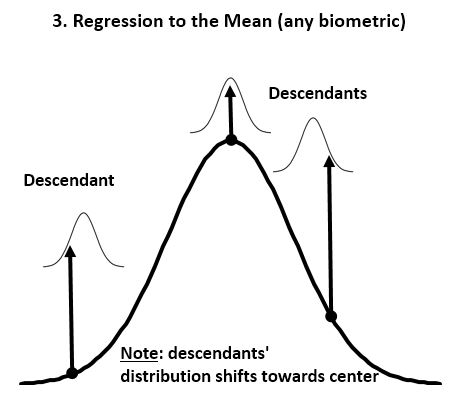
- ‘Regression to the mean’ is the biological law that overrules passive ‘Divergence of Character’. Any homogeneous population can be sorted statistically on various biologic metrics, usually resulting in a Gaussian (normal) distribution that is conserved over time in the absence of major environmental changes (as Mendel first showed; Fig 1&2). ‘Regression to the mean’ is thus the rule that causes the progeny of extreme individuals to be less extreme than their parents. Two outstanding tall parents will have statistically shorter children, and the progeny of the most and least intelligent/strong/aggressive/attractive/etc. will be more average than the parent. Many of the extremes have no descendants at all due to their limitations, and thus their “contribution” to the next generation is simply the average individual.
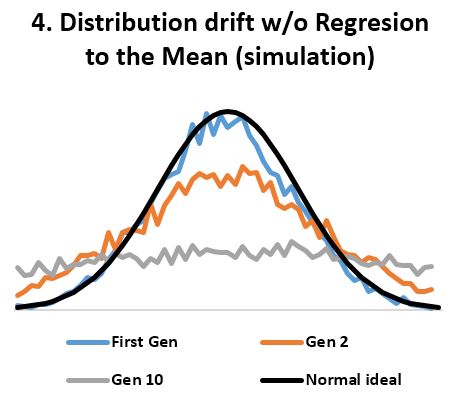
- In stable environments, population variability is extremely well conserved from generation to generation (Fig 3) as documented by the fossil and many other records. ‘Regression to the mean’ is thus a mathematical necessity without which a passive ‘divergence of character’ would be observed in very few generations (Fig 4). ‘Regression to the mean’ mechanism is incredibly accurate and allows for conservation of traits over thousands upon thousands of generations as observed. Scientists were rightfully surprised that ancient bacteria and many other fossils as well as mummified organisms including cats and monkeys are indistinguishable from their contemporary descendants. At a minimum, the number of organisms that show remarkable stability over long periods (living fossils) invalidate the ‘General Divergence’ theory. Does a limited, ‘Special Divergence’ hypothesis still make sense?
- Observed long term regression is highly unexpected and contrary to ‘divergence of character’ and ‘drift’ hypotheses. ‘Regression to the mean’ operates in the longest term observed, whenever environmental conditions are restored following significant changes that led to adaptive mutations. Most – if not all – organisms are endowed with a limited ‘plasticity’ trait that allows them to retain adaptive characteristics for generations. And yet, when the stimulus that caused the adaptation disappears, these organisms regress rather than maintaining those adaptive traits or accumulating even more diverging ones. Darwin’s finches, the peppered moth, antibiotic resistant bacteria and the domesticated plants & animals – all these and more have been observed to regress to the old mean when the adaptive stressor is removed, thus disproving even the limited, ‘Special Divergence’ hypothesis. These are not coincidences! The regression can happen over a few generations as in most epigenetic changes, many generations, and even the indefinite future if the adaptive stimulus is maintained (such as in domestication). Biologic variability can be compared to a loaded spring – the more it stretches, the harder the pull back (regression to the mean) and the more fragile is the extreme variant population. Domesticated plants and animals show that crossbreeds are resilient, while pure breeds are fragile showing that extinction of the extremes is the default outcome that promotes the ‘regression to the mean’ of the extended population.
- Adaptation neither demands not implies divergence in any way. What about the ‘adaptive radiation’ seen in Darwin’s finches, the cichlids of the African Great Lakes, and others? Is this not ‘divergence of character’? No. The driving force in all these and more is adaptation, not divergence even if “evolution” were true. Organisms just seek survival and, if their built-in yet limited plasticity matches the environmental challenges, these populations survive as variants. Otherwise, they simply go extinct like many others before. The new traits are not ‘divergent’ as shown by all known cases of reversals (as discussed) and none of further divergence when the adaptive stressor is removed. If ‘divergence of character’ were true, adaptive plasticity traits would be cumulative and sticky even after the adaptive stressor was removed, and the more extreme variants would be at least as resilient as the mean. Furthermore, experiments would show increasing variability over time in all research organisms and even more so in the short lived ones like bacteria. There would not be any distinct “species” and organisms would freely undergo metamorphosis (transmutation) into one another. Differential survival and randomness would eliminate all but the “best adapted” allele, therefore the Mendelian conservation of alleles would not be observed. Yet none of these are happening, thus falsifying the ‘divergence of character’ hypothesis.
- Adaptation is “fast and done”, “do or die” by necessity, unlike the supposed “slow and ongoing” ‘divergence of character’. If adaptation is not fast enough, the population simply goes extinct as many others did. The cichlids of Lake Victoria had less than 15,000 years to adapt and are as diverse if not more so than the cichlids in the other, much older African Great Lakes. But they do not need even that much time as the newer aquarium varieties obtained in a few generations show. Most likely, cichlids variants have come and gone throughout the history of all African Great Lakes in short cycles of adaptation. And that is why the cichlid biodiversity difference between a few years (Lake Victoria) and millions of years (other African Great Lakes) is unremarkable. The only remarkable fact is that cichlids have a predominantly Gondwanan distribution showing that in 180+ mil years, they did not adapt to ocean living despite their otherwise high adaptability. This clearly shows the limitations of adaptability and makes it an unlikely substitute to ‘divergence of character’. Darwin’s finches, peppered moths, bacteria, and many other also adapt fast or die as observed. And when the stimulus disappears, they revert just as quickly, and later readapt to whatever new stimulus they face or simply die out trying as confirmed. It is a very good thing ‘divergence of character’ is false, or else antibiotic resistant bacteria and other superbugs would have killed mankind by now as “evolution” falsely predicted.
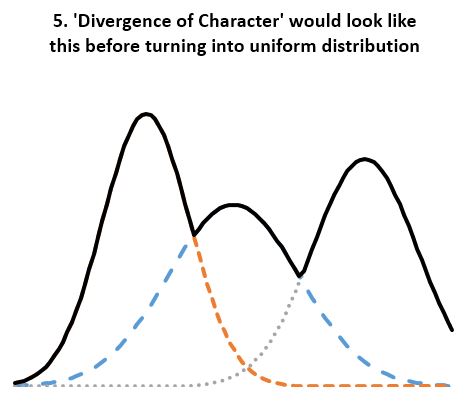
- Statistical evidence refutes ‘divergence of character’. According to the theory, “when organisms compete for scarce resources, natural selection should favor those individuals that are least like their competitors”. And since organisms always “compete for scarce resources”, the least average members of a homogenous population should always be favored by “natural selection”. If so, the well known normal distribution of any organism dimension (length, height, weight, etc.) should always be under pressure to change. We should see groups of “least like” the average form second, third, and so on normal distributions of their own, thus reshaping the original normal distribution into a composite distribution with several peaks and valleys as in Fig 5. And even that should not be adequate, as any concentration of similar individuals would be disadvantaged according to the ‘Divergence of character’ hypothesis, thus leading to uniform distributions as in Fig 4. However, neither Fig 5 nor uniform distributions are seen in homogeneous populations. Instead, we always see normal distributions. And since we see the normal distribution maintained over arbitrary number of generations and no hint of transitioning to a uniform distribution, the ‘Divergence of character’ hypothesis must be discarded. A trend not supported by several period observations must be discarded as noise artifact. This is the case for all examples considered including Darwin’s finches, the peppered moth, antibiotic resistant bacteria, cichlids, etc. All seem somewhat supportive of the divergence hypothesis over carefully chosen periods, yet the divergence is clearly illusory over longer periods.
- Are the bear of North America not like Fig 5? Yes, but they occupy different geographic regions. They are not homogenous. Indeed, we do encounter subfamilies of organisms, that have normally distributed metrics within the subgroup yet clearly distinct from those of other subgroups. However, where these subgroups overlap, the blend is always geographic and never biologic, meaning we see fewer of one kind and more of the other when moving from one’s territory to the others’ instead of blended characteristics as ‘divergence of character’ would predict. Humans are not different “species” although various subgroups are exclusively vegan/carnivorous, white/black, extra small/large. And domesticated organisms including canids are even more diverse than humans. Are the wild cichlids, finches, mice, and others qualitatively different than humans and canids? No. Then why the different “species”, many of which, ironically, are threatened by hybridization? The unwarranted inflation of “species” that do not even meet the loosest definition of reproductive isolation has the sole purpose of perpetuating the myth of ‘divergence of character’.
- Multimodal distributions in homogenous populations are not due to ‘divergence of character’. Indeed, bimodal distributions (Fig 2) and multimodal distributions are not uncommon in homogenous populations. However, these are due to the discreteness of physics in general and biology in particular, not due to ‘divergence of character’. Male and female populations are not diverging from one another and various alleles are in long term cyclical equilibrium as shown (spring model). ‘Drift’ is often invoked as a mechanism of ‘divergence of character’. This is wrong because ‘drift’ explains nothing as it is either aimless noise or due to adaptation and environmental change. Yet, as shown, adaptation is in no way ‘divergence of character’. In addition, the stable coexistence of several distinct variants within a homogenous population shows “gradualism”, “survival of the fittest”, and “natural selection” to be false because the alleles responsible are themselves distinct (no “gradualism”), they all “survive”, and neither is “selected” for or against.
- Darwin worried about regression to the mean for the wrong reasons. Namely, if blending inheritance (Darwin laid an egg) was true, then natural selection could not be true. Darwin puzzled over this a lot, but ended up with nothing satisfactory. Then Mendel showed that inheritance is discrete, not blended. Mendelian Inheritance Tables (see Punnett squares / Hardy-Weinberg equilibrium) show “probabilistic traits conservation” and thus disproving ‘divergence of character’ (at least as byproduct of reproduction) as well as dismissing “gradualism” (another one of Darwin’s unsupported claims).
- When entire populations split, do subgroups diverge from one another? This is not how ‘divergence of character’ is supposed to work.Descendants are supposed to diversify within the homogenous population. Furthermore, populations split by environmental conditions simply adapt to the new environment and for as long as those conditions allow. Adaptation is the driving force with no ‘divergence of character’ anywhere in sight. Island biology is the most diverse because islands are isolated and have many microenvironments. However, island variants are close descendants of their original colonists, showing that no divergence ever happened. Their risk of hybridization is high, disproving the “speciation” claim. They are also fragile examples of the extreme stretched biological spring model discussed, and will likely go extinct if at all stressed and when interacting with mainland.
Summary:
1. ‘Regression to the mean’ is the biological law that overrules passive ‘Divergence of Character’
2. In stable environments, population variability is extremely well conserved from generation to generation
3. Observed long term regression is highly unexpected and contrary to ‘divergence of character’ and ‘drift’ hypotheses
4. Adaptation neither demands not implies ‘divergence of character’ in any way
5. Adaptation is “fast and done”, “do or die” by necessity, unlike the supposed “slow and ongoing” ‘divergence of character’
6. Adaptation has limited powers and is thus not a substitute for ‘divergence of character’
7. ‘Divergence of character’ hypothesis would lead to uniform rather than normal (Gaussian) distributions as observed in homogenous populations
8. A trend not supported by several period observations must be discarded as noise artifact
9. The unwarranted inflation of “species” that do not even meet the loosest definition of reproductive isolation has the sole purpose of perpetuating the myth of ‘divergence of character’
10. Multimodal distributions in homogenous populations are not due to ‘divergence of character’
11. Mendelian tables show “probabilistic traits conservation”, disproving ‘divergence of character’ (at least as byproduct of reproduction), as well as dismissing ‘gradualism’
12. Island biology proves adaptation and the biologic spring model while disproving ‘divergence of character’
13. What’s in, what’s out? IN: ‘regression to the mean’, ‘adaptation’, coexisting variants, long term stability, spring model, normal distributions. OUT: ‘divergence of character’, gradualism, drift, speciation, uniform distributions, “natural selection”, “survival of the fittest”, “evolution”.
Links:
https://ucmp.berkeley.edu/bacteria/bacteriafr.html
https://www.sciencedaily.com/releases/2019/10/191018112136.htm
https://www.ncbi.nlm.nih.gov/pmc/articles/PMC3285564/
https://www.ncbi.nlm.nih.gov/pmc/articles/PMC3352989/
http://www.galton.org/essays/1880-1889/galton-1886-jaigi-regression-stature.pdf
https://en.wikipedia.org/wiki/Character_displacement
https://en.wikipedia.org/wiki/Sympatric_speciation
https://www.bionity.com/en/encyclopedia/Character_displacement.html
https://biologydictionary.net/divergent-evolution/
https://en.wikipedia.org/wiki/Cichlid
https://biology.stackexchange.com/questions/41982/regression-to-the-mean-and-evolution
Regression to the mean is something we observe in biparental reproduction. I am using the phrase as Galton and Pearson used it, as describing a single generation of reproduction, not with regard to long-term change, as nonlin.org is the only person I have seen use it for that.
If you observe individuals, don’t see who they mate with, and ask about the mean of their offspring, you will find under many schemes of mating, including random mating, that an tall individual has offspring that are on average smaller than it is. And a small individual has offspring that are on average taller than it is. This is true even with perfect heritability of height. Why does it occur? Because these extreme individuals on average mate with less extreme individuals.
What is wrong with nonlin.org’s use of clonal reproduction as an example is that when we measure the parent, it will be like measuring both parents when there is random mating. But observations of regression to the mean measure how the mean of the offspring is related to the phenotype of one of the two parents.
Moi? Au contraire, it’s you. Look above. You have implied differential survival between phenotypes twice in examples that are supposed to demonstrate “regression to the mean” as a conservative mechanism. You just keep on invoking selection and fitness, when all I am asking for is that you show some support for the existence of long-term regression to the mean. If you don’t like fitness as a concept, then stop making an appeal to it.
Once more:
Heehee. Are your ancestors still alive? Send my regards to them, will you?
Of course the ancestors of horses are not alive. They would have to be very, very, very old, wouldn’t they? But have it your way: which contemporary species of eohippus do you believe to be the ancestors of modern horses? Then, we can start discussing whether horses are currently in their original form.
Wow, is there no established fact that you will not deny? The increase in antibiotic resistance is very real and a major health concern.
So what well established fact will you be denying next? The sky isn’t blue? The sun isn’t warm? Water isn’t wet?
Apparently not.
Your comments denote Zero understanding of what goes on. I very much doubt a discussion with someone that understands Nothing makes any sense.
However, at least you finally replied instead of just talking to yourself, so…
1. How can you say that? Don’t you see the rand() function in the formula? Is that not introducing noise?
2. Regression, yes. Your “why” is FALSE. Because of 6. and 7. below.
3. Look at the formulas again. What “not really”? Yes or No? The child’s distribution is normal and centered on the parent. That means “no regression”. Really.
5. Explain. Examples from nature?
6. FALSE. Can you have divergence without spread or spread without divergence? NO WAY. Examples from nature?
7. You can do whatever in a model, no doubt. I added zero regression and you want to add x%. Who cares? The real question is: is your model realistic? NO. Why? Because the populations distributions remain CONSTANT over the observable horizon. They do not converge or diverge as in your model.
That’s why your 6. is false. And this looks like the fundamental disagreement. Why did it take you so long to clarify something as simple as this?
Think on this, non-lin. All modellers should bear this in mind. Including you.
I answered your questions. You owe me the courtesy of answering mine as asked. Will you try again? a.b.c.d.?
We established that “random mating” is false for many/most organisms. Will you acknowledge?
Mating with less extremes is only a partial reason (regression a). Most extremes simply don’t mate because no one wants them (regression b). As such, their “legacy” is just the average (like a voter that doesn’t vote). There’s also an intrinsic regression well illustrated by the Curie family (regression c) https://en.wikipedia.org/wiki/Curie_family. There may be other regression reasons.
Huh?
We discussed this. “Differential survival” is a meaningless soundbite. We’re all different and some survive. None of those are what you think. You’re like the drunk that sees a party everywhere. The environment does matter of course. Yet “divergence of character” is supposed to happen in a homogeneous population that experiences the same constant environment. That’s no “environmental differential” to you.
Long term reversion is quite clear. Think about it: why would all those organisms go back? Why not in a random other direction?
Forget horses, ancestors, and anything else that cannot be double and triple checked. Science has to be strictly verified experimentally. And there’s plenty organisms available. Heck, even humans can be reliably traced (double and triple checked) 250 generations (that’s Otzi and the mummies). My model showed effects in 10 gen, so you have no valid reason to appeal to hocus-pocus millions and billions of years. Like I said: “A trend not supported by several period observations must be discarded as noise artifact.” Even mountain rising can be observed here and now as discussed. And you have yet to come up with a counterexample on that abandoned discussion. Sometimes you seem so amazingly logic-proof.
Yet “naturally selected” and “higher fitness” AB-resistant bacteria have not taken over the world. WHY NOT? Because they’re not “evolved”. Because they have not suffered any “beneficial mutations”. That’s why. And how do we fight these monsters? By REDUCING the use of antibiotics! Does this make any “evolutionary” sense to you?
Are you writing back to me something I thought and said?!? What’s wrong with you?
nonlin.org: Regression toward the mean is a standard topic in quantitative genetics, so I don’t care about your particular questions, I want to see if you understand why the quantitative genetics observation that the average of offspring of one parent are in general closer to the population mean than is the parent itself. The observation is based on looking at one parent, who mates with one or more others that are not observed.
The phenomenon occurs because of the effect of noninherited environmental effects or dominance interactions that are not inherited. It also occurs because of the contribution of the other mate.
It occurs in all the simplest models, for these obvious reasons. If you want to argue that these models are no good, you are welcome to, and I will ignore what you write.
So you do not know and believe it is impossible to know where the modern horse comes from?
You have put forward several arguments that rely on the fact that survival and reproduction of individuals is non-random with respect to phenotype. When you feel compelled to deny facts that your own arguments rely on, you are doing something wrong.
If you want to pretend that what you are doing is science, then demonstrate how you “strictly verified” your own claims. So far, you have not told us how you know that many species are currently not in their original form. Denying that the fossil record could be helpful in this matter is not really helping your case either.
Because antibiotics have only been in widespread use for several decades.
Yep. Also consider that bacteria didn’t take over the world before the use of antibiotics either. They lived in an equilibrium with their hosts. Sometimes killing whole populations, but not the entire population, sometimes just making them sick for some periods of time, sometimes killing their hosts slowly, etc. That’s where we could go.
What’s happening is that as antibiotic resistance increases so will death by infection. Instead of cancer and heart disease, we’ll have infections back at the top of the list. We’re not there because there’s been development of new antibiotics, and because antibiotic resistance is not universal towards every antibiotic, and it’s not universal across all bacteria.
Not every bacteria is a pathogen, and not every pathogen is a mortal pathogen.
Nothing to do with “regression to the mean.”
I don’t care one bit about “your” models. I wrote something clear, concise and very well supported. You only refuse to answer because you know I’m 100% right. So long.
Not what I said. When you tell me it comes from XYZ but we see ABSOLUTELY no change in thousands of generations of horses that we can double and triple check, you’re blowing smoke. IOW, recent, provable history doesn’t fit your claim.
You’re misrepresenting and erroneously extrapolating what I said.
Done. Read.
I cited a number of examples which we know experimentally to transition on stimulus and regress on stimulus removal. It’s improper to speak of anything that cannot be verified experimentally.
The burden of proof is on you – not on me – and you’re not meeting it:
“Several decades” represent millions if not billions of bacteria generations. Plenty of time to see a trend. Which we DON’T see. And in fact we see THE OPPOSITE. Cut down antibiotic use and ab-resistant bacteria disappear. Now, that’s inconvenient science!
FALSE. We’re not there because we’re reducing the use of antibiotics. We will never be “there”, now that we smartened up.
Of course it’s a regression. The bacteria population doesn’t disappear but it’s losing its ab-resistant properties. Goes to show ab-resistance is not “evolution”. No “fitness” and no “beneficial mutation” either.
I wasn’t talking to you because you’re uneducable. Yet: what I said is true: antibiotic resistance is not universal either in antibiotic scope, or in presence across bacteria. It’s also true that bacteria did not “conquer the world” before the use of antibiotics. Why did you ignore this part?
You think it’s just because “we’ve smartened up.” Well, I hope you’re right that we’ve smartened up, because in most of the countries I visit the “smartening up” does not seem to be happening. In those where it is imagined to be happening it doesn’t seem to go much beyond political pronunciations.
Of course it isn’t.
That’s what we hope will happen once we stop. This is based on evolutionary principles, not on some poorly understood magical mathematical model about “regressing” towards some imaginary “mean.”
Goes to show that you do not understand evolution one bit. Evolution is not a claim that adaptations never disappear. The worse of it is that you’re holding to evolutionary phenomena to deny evolution. Your “argument” is therefore self-refuting. Fitness is relative to the environment. If there’s antibiotics, then fitness means resistance to antibiotics. If there’s no antibiotics, then resistance to antibiotics is no longer part of the fitness landscape of the organisms. While antibiotics are introduced, the antibiotics act as positive selection, whereby organisms with mutations or acquired genes that help them survive, survive. After survival, negative selection, death of the mutants who lose their resistance, keeps the population resisting antibiotics. Both types of selection are evolutionary concepts. If the antibiotic challenge goes away, then no more negative selection, the resistance goes away by drift. Another evolutionary concept. The whole thing you’re describing is evolution, you just want to call it “regression to the mean” because you’re an idiot. That’s all.
I pretty sure that you won’t show understanding of what I wrote (others seems to have made the very same point, and you missed it). So I’ll go back to ignoring your obtuse “answers.”
Here’s how one can attempt to understand nonlin’s delightfully incoherent idea-set.
[Warning: do not attempt without adequate safety gear.]
Forget everything you know about biology. Forget about genotype-phenotype mapping, mutation, selection, mating (whether assortative or not), haploids, diploids, dominance etc. Forget biochemistry. Forget about the debate over regression to the mean and blended inheritance.
All this is irrelevant.
All you need to know is that nonlin once heard that “regression to the mean” presented a problem for Darwin, and that he also heard some out-of-context comments about Galapagos finch beak sizes and antibiotic resistance, which he has woefully misinterpreted.
This is nonlin’s model, in essence:
Consider a box in outer space, inside it particles undergo bounded random walks. (Note the lack of reproduction here; he doesn’t need to model it.)
At steady state, the distribution in the box will be uniform, and the average position will be slap bang in the middle of the box. [Remember his stuff about uniform distributions and ‘hard natural limits’? Oh yeah.]
Now imagine that a ‘selective’ force is applied to the particles, such that moving leftwards is more likely than moving rightwards — a nearby planet would do the trick. The particles will arrive at a new steady state, where the particles are unevenly distributed with a mean somewhat to the left.
Next, remove the ‘selective’ force, and the particles will revert to the original, uniform distribution. Nonlin is confusing this reversion in the absence of ‘selective force’ with ‘regression to the mean’, which (as Joe has repeatedly pointed out) is always regression to the NEW mean, wherever that might be.
It’s got nothing to do with biology; nonlin believes that there is an inconsistency between evolutionary theory and observed stasis (over a few dozen generations).
What he doesn’t understand is that, over evolutionary timescales, the edges of his box may move
We’ve ALL been over-thinking this.
It’s become a recurring pattern.
.
.
It’s work in progress. But one thing is clear: if ab-resistant bacteria were “evolved” and more “fit”, you would not fight it by reducing the use of antibiotics.
FALSE. As explained above. Cutting down on antibiotics is not “based on” but instead it goes AGAINST “evolutionary principles”.
So if I have wings then that’s fitness” and if I don’t have wings then that’s also “fitness”. Goes to show the whole thing is crap.
Whatever “explains everything”, explains nothing. To not mention the nonexistent predictive power.
FALSE. There is no “NEW mean” in thousands and millions of generations as observed. There is no “evolutionary timescales”. Only bullshit behind that curtain. If something is truly happening over the long therm, then we MUST observe a fraction of it happening over the short term. We DO see mountains rising TODAY even though they reach their peaks over geological timescales.
As explained:
“And since we see the normal distribution maintained over arbitrary number of generations and no hint of transitioning to a uniform distribution, the ‘Divergence of character’ hypothesis must be discarded. A trend not supported by several period observations must be discarded as noise artifact. This is the case for all examples considered including Darwin’s finches, the peppered moth, antibiotic resistant bacteria, cichlids, etc. All seem somewhat supportive of the divergence hypothesis over carefully chosen periods, yet the divergence is clearly illusory over longer periods.”
Nonlin.org,
As I already said:
You, as the good sport that you are, showed no understanding of what I wrote, and I’m back to ignoring your non-answers.
P.S.
Have you finished elementary school yet? Reading comprehension is a must to pass the literacy test. I know that a lot of people pass it regardless of being almost as illiterate as yourself, but you should not risk it. Here’s the appropriate level for you to get started.
Don’t try and have a conversation with me until you’ve passed those literacy tests and showed some actual reading for comprehension abilities. Until then, you should stop pretending that you’re above preschool.