
As shown repeatedly, “Nothing in Evolution Makes Sense. Period.” Not natural selection, gradualism, human evolution, UCD, tree of life, etc. And just to confirm, let’s look at another one of the nonsensical concepts of “evolution”.

- ‘Divergence of character’ (character displacement or sympatric speciation) postulates: “during the incessant struggle of all species to increase in numbers, the more diversified these descendants become, the better will be their chance of succeeding in the battle of life. Thus the small differences distinguishing varieties of the same species, will steadily tend to increase till they come to equal the greater differences between species of the same genus, or even of distinct genera” (Darwin 1859). Sympatric speciation is hypothesized as “the evolution of a new species from a surviving ancestral species while both continue to inhabit the same geographic region”.
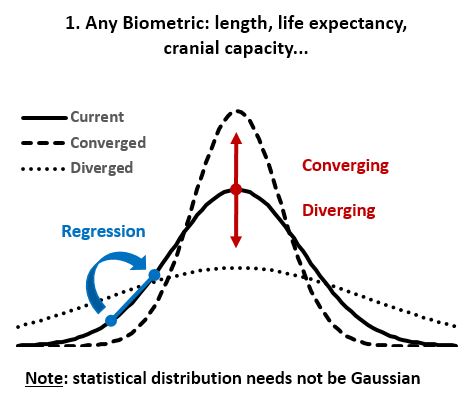
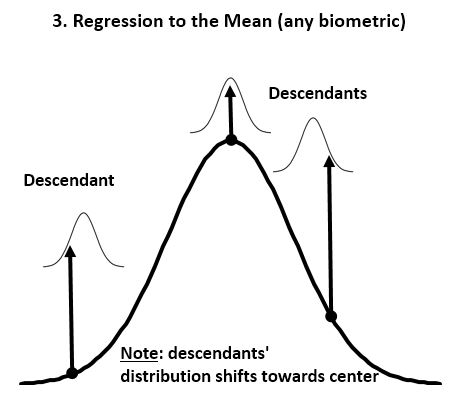
- ‘Regression to the mean’ is the biological law that overrules passive ‘Divergence of Character’. Any homogeneous population can be sorted statistically on various biologic metrics, usually resulting in a Gaussian (normal) distribution that is conserved over time in the absence of major environmental changes (as Mendel first showed; Fig 1&2). ‘Regression to the mean’ is thus the rule that causes the progeny of extreme individuals to be less extreme than their parents. Two outstanding tall parents will have statistically shorter children, and the progeny of the most and least intelligent/strong/aggressive/attractive/etc. will be more average than the parent. Many of the extremes have no descendants at all due to their limitations, and thus their “contribution” to the next generation is simply the average individual.
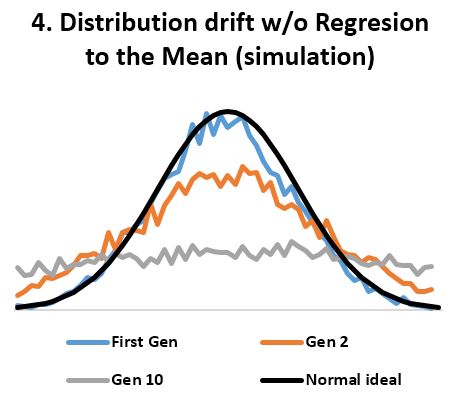
- In stable environments, population variability is extremely well conserved from generation to generation (Fig 3) as documented by the fossil and many other records. ‘Regression to the mean’ is thus a mathematical necessity without which a passive ‘divergence of character’ would be observed in very few generations (Fig 4). ‘Regression to the mean’ mechanism is incredibly accurate and allows for conservation of traits over thousands upon thousands of generations as observed. Scientists were rightfully surprised that ancient bacteria and many other fossils as well as mummified organisms including cats and monkeys are indistinguishable from their contemporary descendants. At a minimum, the number of organisms that show remarkable stability over long periods (living fossils) invalidate the ‘General Divergence’ theory. Does a limited, ‘Special Divergence’ hypothesis still make sense?
- Observed long term regression is highly unexpected and contrary to ‘divergence of character’ and ‘drift’ hypotheses. ‘Regression to the mean’ operates in the longest term observed, whenever environmental conditions are restored following significant changes that led to adaptive mutations. Most – if not all – organisms are endowed with a limited ‘plasticity’ trait that allows them to retain adaptive characteristics for generations. And yet, when the stimulus that caused the adaptation disappears, these organisms regress rather than maintaining those adaptive traits or accumulating even more diverging ones. Darwin’s finches, the peppered moth, antibiotic resistant bacteria and the domesticated plants & animals – all these and more have been observed to regress to the old mean when the adaptive stressor is removed, thus disproving even the limited, ‘Special Divergence’ hypothesis. These are not coincidences! The regression can happen over a few generations as in most epigenetic changes, many generations, and even the indefinite future if the adaptive stimulus is maintained (such as in domestication). Biologic variability can be compared to a loaded spring – the more it stretches, the harder the pull back (regression to the mean) and the more fragile is the extreme variant population. Domesticated plants and animals show that crossbreeds are resilient, while pure breeds are fragile showing that extinction of the extremes is the default outcome that promotes the ‘regression to the mean’ of the extended population.
- Adaptation neither demands not implies divergence in any way. What about the ‘adaptive radiation’ seen in Darwin’s finches, the cichlids of the African Great Lakes, and others? Is this not ‘divergence of character’? No. The driving force in all these and more is adaptation, not divergence even if “evolution” were true. Organisms just seek survival and, if their built-in yet limited plasticity matches the environmental challenges, these populations survive as variants. Otherwise, they simply go extinct like many others before. The new traits are not ‘divergent’ as shown by all known cases of reversals (as discussed) and none of further divergence when the adaptive stressor is removed. If ‘divergence of character’ were true, adaptive plasticity traits would be cumulative and sticky even after the adaptive stressor was removed, and the more extreme variants would be at least as resilient as the mean. Furthermore, experiments would show increasing variability over time in all research organisms and even more so in the short lived ones like bacteria. There would not be any distinct “species” and organisms would freely undergo metamorphosis (transmutation) into one another. Differential survival and randomness would eliminate all but the “best adapted” allele, therefore the Mendelian conservation of alleles would not be observed. Yet none of these are happening, thus falsifying the ‘divergence of character’ hypothesis.
- Adaptation is “fast and done”, “do or die” by necessity, unlike the supposed “slow and ongoing” ‘divergence of character’. If adaptation is not fast enough, the population simply goes extinct as many others did. The cichlids of Lake Victoria had less than 15,000 years to adapt and are as diverse if not more so than the cichlids in the other, much older African Great Lakes. But they do not need even that much time as the newer aquarium varieties obtained in a few generations show. Most likely, cichlids variants have come and gone throughout the history of all African Great Lakes in short cycles of adaptation. And that is why the cichlid biodiversity difference between a few years (Lake Victoria) and millions of years (other African Great Lakes) is unremarkable. The only remarkable fact is that cichlids have a predominantly Gondwanan distribution showing that in 180+ mil years, they did not adapt to ocean living despite their otherwise high adaptability. This clearly shows the limitations of adaptability and makes it an unlikely substitute to ‘divergence of character’. Darwin’s finches, peppered moths, bacteria, and many other also adapt fast or die as observed. And when the stimulus disappears, they revert just as quickly, and later readapt to whatever new stimulus they face or simply die out trying as confirmed. It is a very good thing ‘divergence of character’ is false, or else antibiotic resistant bacteria and other superbugs would have killed mankind by now as “evolution” falsely predicted.
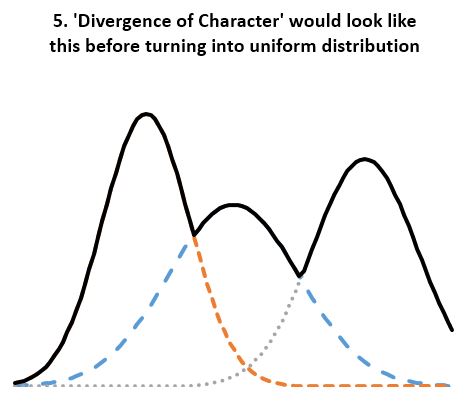
- Statistical evidence refutes ‘divergence of character’. According to the theory, “when organisms compete for scarce resources, natural selection should favor those individuals that are least like their competitors”. And since organisms always “compete for scarce resources”, the least average members of a homogenous population should always be favored by “natural selection”. If so, the well known normal distribution of any organism dimension (length, height, weight, etc.) should always be under pressure to change. We should see groups of “least like” the average form second, third, and so on normal distributions of their own, thus reshaping the original normal distribution into a composite distribution with several peaks and valleys as in Fig 5. And even that should not be adequate, as any concentration of similar individuals would be disadvantaged according to the ‘Divergence of character’ hypothesis, thus leading to uniform distributions as in Fig 4. However, neither Fig 5 nor uniform distributions are seen in homogeneous populations. Instead, we always see normal distributions. And since we see the normal distribution maintained over arbitrary number of generations and no hint of transitioning to a uniform distribution, the ‘Divergence of character’ hypothesis must be discarded. A trend not supported by several period observations must be discarded as noise artifact. This is the case for all examples considered including Darwin’s finches, the peppered moth, antibiotic resistant bacteria, cichlids, etc. All seem somewhat supportive of the divergence hypothesis over carefully chosen periods, yet the divergence is clearly illusory over longer periods.
- Are the bear of North America not like Fig 5? Yes, but they occupy different geographic regions. They are not homogenous. Indeed, we do encounter subfamilies of organisms, that have normally distributed metrics within the subgroup yet clearly distinct from those of other subgroups. However, where these subgroups overlap, the blend is always geographic and never biologic, meaning we see fewer of one kind and more of the other when moving from one’s territory to the others’ instead of blended characteristics as ‘divergence of character’ would predict. Humans are not different “species” although various subgroups are exclusively vegan/carnivorous, white/black, extra small/large. And domesticated organisms including canids are even more diverse than humans. Are the wild cichlids, finches, mice, and others qualitatively different than humans and canids? No. Then why the different “species”, many of which, ironically, are threatened by hybridization? The unwarranted inflation of “species” that do not even meet the loosest definition of reproductive isolation has the sole purpose of perpetuating the myth of ‘divergence of character’.
- Multimodal distributions in homogenous populations are not due to ‘divergence of character’. Indeed, bimodal distributions (Fig 2) and multimodal distributions are not uncommon in homogenous populations. However, these are due to the discreteness of physics in general and biology in particular, not due to ‘divergence of character’. Male and female populations are not diverging from one another and various alleles are in long term cyclical equilibrium as shown (spring model). ‘Drift’ is often invoked as a mechanism of ‘divergence of character’. This is wrong because ‘drift’ explains nothing as it is either aimless noise or due to adaptation and environmental change. Yet, as shown, adaptation is in no way ‘divergence of character’. In addition, the stable coexistence of several distinct variants within a homogenous population shows “gradualism”, “survival of the fittest”, and “natural selection” to be false because the alleles responsible are themselves distinct (no “gradualism”), they all “survive”, and neither is “selected” for or against.
- Darwin worried about regression to the mean for the wrong reasons. Namely, if blending inheritance (Darwin laid an egg) was true, then natural selection could not be true. Darwin puzzled over this a lot, but ended up with nothing satisfactory. Then Mendel showed that inheritance is discrete, not blended. Mendelian Inheritance Tables (see Punnett squares / Hardy-Weinberg equilibrium) show “probabilistic traits conservation” and thus disproving ‘divergence of character’ (at least as byproduct of reproduction) as well as dismissing “gradualism” (another one of Darwin’s unsupported claims).
- When entire populations split, do subgroups diverge from one another? This is not how ‘divergence of character’ is supposed to work.Descendants are supposed to diversify within the homogenous population. Furthermore, populations split by environmental conditions simply adapt to the new environment and for as long as those conditions allow. Adaptation is the driving force with no ‘divergence of character’ anywhere in sight. Island biology is the most diverse because islands are isolated and have many microenvironments. However, island variants are close descendants of their original colonists, showing that no divergence ever happened. Their risk of hybridization is high, disproving the “speciation” claim. They are also fragile examples of the extreme stretched biological spring model discussed, and will likely go extinct if at all stressed and when interacting with mainland.
Summary:
1. ‘Regression to the mean’ is the biological law that overrules passive ‘Divergence of Character’
2. In stable environments, population variability is extremely well conserved from generation to generation
3. Observed long term regression is highly unexpected and contrary to ‘divergence of character’ and ‘drift’ hypotheses
4. Adaptation neither demands not implies ‘divergence of character’ in any way
5. Adaptation is “fast and done”, “do or die” by necessity, unlike the supposed “slow and ongoing” ‘divergence of character’
6. Adaptation has limited powers and is thus not a substitute for ‘divergence of character’
7. ‘Divergence of character’ hypothesis would lead to uniform rather than normal (Gaussian) distributions as observed in homogenous populations
8. A trend not supported by several period observations must be discarded as noise artifact
9. The unwarranted inflation of “species” that do not even meet the loosest definition of reproductive isolation has the sole purpose of perpetuating the myth of ‘divergence of character’
10. Multimodal distributions in homogenous populations are not due to ‘divergence of character’
11. Mendelian tables show “probabilistic traits conservation”, disproving ‘divergence of character’ (at least as byproduct of reproduction), as well as dismissing ‘gradualism’
12. Island biology proves adaptation and the biologic spring model while disproving ‘divergence of character’
13. What’s in, what’s out? IN: ‘regression to the mean’, ‘adaptation’, coexisting variants, long term stability, spring model, normal distributions. OUT: ‘divergence of character’, gradualism, drift, speciation, uniform distributions, “natural selection”, “survival of the fittest”, “evolution”.
Links:
https://ucmp.berkeley.edu/bacteria/bacteriafr.html
https://www.sciencedaily.com/releases/2019/10/191018112136.htm
https://www.ncbi.nlm.nih.gov/pmc/articles/PMC3285564/
https://www.ncbi.nlm.nih.gov/pmc/articles/PMC3352989/
http://www.galton.org/essays/1880-1889/galton-1886-jaigi-regression-stature.pdf
https://en.wikipedia.org/wiki/Character_displacement
https://en.wikipedia.org/wiki/Sympatric_speciation
https://www.bionity.com/en/encyclopedia/Character_displacement.html
https://biologydictionary.net/divergent-evolution/
https://en.wikipedia.org/wiki/Cichlid
https://biology.stackexchange.com/questions/41982/regression-to-the-mean-and-evolution
Alan Fox,
Or maybe its like a fat hippopotamus sculpture?
phoodoo,
The pink dress metaphor works for me.
Right, the idea that YOU think belief in religion is a reproductive advantage and that’s how it emerged, is just like someone sees a pink dress and someone sees a gold one.
Exactly like that. Exactly. Except not.
No. I may put up an OP.
ETA: Link
This is just ignorant nonsense.
Can’t link the two. You know and accept regression to the mean, so I need not explain it to you. It’s an observable fact if not fully understood. It applies to populations, not to individuals. If you have ANY doubts, just look at all good and bad outstanding individuals in ANY domain and note that none of their descendants has been more outstanding then themselves over a few generations.
To contrast, I explained with logic why “fitness” is just bullshit. So your position is untenable. Either:
1. reply with your “fitness” function,
2. admit “fitness” is bullshit, OR
3. come up with a VERY GOOD reason why you can’t quantify something this important
Enough stalling. Enough bullshit.
Not the Darwinist “selection” for sure. Here we have a big, immutable anchor whereas the Darwinist myth specifically claims “no anchor”. Once again, I know your myths better than you do. Are you still not embarrassed? And that aside from “natural selection” being as stupid of a concept as “arising”.
I am not sure what you’re asking. You have the formula: it’s just a full size random normal centered on the parent. What would you change? Obviously, offset and/or skewness are responsible for regression, so I cannot change (add) them since I wanted to show an universe WITHOUT regression.
The “parent” represents both. 50/50.
The formula in cell B3 begins “=A3+(SQRT(…”
Change it to “=0.9*A3+(SQRT(…” , so that each offspring receives 90% of the parental ‘value’, instead of 100%.
In this case, would your model then show
A) “divergence of character”
B) regression to the mean
C) both
?
Also, would this modification make your model more realistic, or less?
Obviously…
No knowledgeable person would make that claim either. But it seems about par for your posts – that we must pretend reality doesn’t exist to even BEGIN to discuss anything with you, and that those who believe in reality are spouting ignorant nonsense. With ground rules like that, why discuss anything?
I just won’t for the same reasons that you don’t want to calculate your regression to the mean: it’s a hassle without any clear benefit.
Put in some effort and I may be tempted to do likewise. Otherwise: find yourself a book on population genetics or life history theory and educate yourself.
Yes, the “Darwinist” selection for sure: Differential survival and reproduction correlated with a phenotypic trait.
Every individual in your spreadsheet has ONE (= 1) parent. A mommy and a daddy makes two. This is as basic as it gets, Nonlin.
Well, that’s an offset to the center, right? AKA ‘regression to the mean’ which I specifically wanted to remove to see what happens in its absence. More importantly, I proved what I wanted to prove. If you want to show something else, be my guest.
So you just chose to be delusional. Good for you.
Ditto.
Nothing stops you from taking the average of the two parents. To not say anything of bacteria. Why are you such a xenophobe, misogynist, and bacteriophobe?
So each “parent” value is obtained by picking two parents at random and averaging their trait values? I bet not, but it should be done that way, at least for a trait that is totally determined by additive genetic effects with no dominance deviations or environmental effects.
Oops, I was wrong. Better make that, for the case where all the variation is heritable (all of it is due to additive genetic variation), we take the average of two randomly chosen parents, then add a normally distributed amount that has mean zero and variance equal to 1/2 the phenotypic variance of the parents.
Nonlin.org,
Your comment appears to have landed in guano. I suggest you repost it without the “delusional” stuff. And your last sentence my need a 🙂 . I fear that was misread as an insult.
To pre-emptively reply to your suggestion to take the average of both parents: Please see Joe’s comment above. If you just take the average, you miss out on the variation caused by Mendelian segregation.
Don’t you just love genetics?
I have an even better idea. He should just repost, and pretend he is an atheist. It’s sure to be fine then.
In a similar way to the way you pretend to be a theist you mean? A theist too cowardly to explain what they actually believe in? Heard that cock crow three times yet phoodoo?
Corneel,
I’ve restored the comment. Apologies for the inconvenience. I overlooked the possibility of humour.
Who would’ve guessed that even creationists themselves are bound by Poe’s law?
You sure do.
Ah, so that’s what you meant by an “offset”. Thank you for the clarification.
I can understand how you might consider that a way of “unfairly” introducing “regression to the mean”, but I was only trying to keep the calculations within your apparent capabilities.
You are still wrong about ‘regression to the mean’.
Here’s a slightly more complicated way of illustrating this:
Now, does this new array show
A) “divergence of character”
B) regression to the mean
C) both
?
Also, would this modification make your model more realistic, or less?
First quote was from OM not Alan.
newton,
No kidding. You are a Svengali.
I wonder why I put it under Alan? Maybe it was because it was only one post before that one, and Alan conveniently overlooked it. Hmm, who knows. Now maybe he will notice it and still ignore it.
No, no, not that. I just thought it was interesting, and oh, wait, are we going to get banned for this? Wait, what are we talking about? I am just talking about divergence of character. I have no idea what you are talking about. Something you don’t give a shit about I think.
That makes no sense , you should still attribute the comment who made it.
Maybe but your attacking me by calling me a Svengali is deeply hurtful. I didn’t imply you were chicken to present your theistic views. I think that is your business.
Most likely guano and scolding,
Sounds like moderation. I don’t but that doesn’t make it less fun to correct you.
It’s still not clear what you want to say.
How would you have modeled it differently?
What’s “randomly chosen parents”?
What’s “1/2 the phenotypic variance of the parents”?
Perhaps you misunderstand the purpose of my model: not an accurate representation of nature (a monumental task), but a simple model to see what happens if ‘regression to the mean’ is disregarded.
Ditto.
Yes, let me rephrase: if you think anyone gives a fuck about your “benefit” in the narrow sense of winning a stupid argument, you’re insane. How’s that? And here it is: 🙂
I should have added to the list http://theskepticalzone.com/wp/divergence-of-character-myth/comment-page-8/#comment-268034
4. plead insanity
But then again, the insanity defense is indeed “VERY GOOD reason” so 3. works just as well.
You tell me. I was clear enough: if you want to show something, you do it. And I’ll be happy to critique it.
Oh dear.
You appear to be unable to make a simple edit to an Excel model. Is this model actually yours, or did you ‘borrow’ it from someone else? Would you care to explain why you used the Box-Muller instead of the NORMINV()?
I guess my fantasy of helping you educate yourself will remain just that, a fantasy.
Oh well.
Your model, with the introduction of noise, becomes a lot more realistic. It also shows (pay attention here) regression to the mean and “divergence of character. At the same time.
The hilarious bit here is that other posters are trying to help you make your model more realistic in a bunch of more complicated and challenging ways. Fat chance, if you cannot manage a very minor edit.
Remember, nonlin, your ‘argument’ has the following form :
A) I, nonlin, have built a really bad model.
B) It shows behavior X because it lacks effect Y.
C) This is not seen in nature, therefore effect Y holds throughout nature.
Various posters have explained to you how only statement A is true.
I was using standard terminology from quantitative genetics. That is the theory that is used by animal and plant breeders when they set up breeding systems. They have millions of dollars at stake so they have to know their stuff. You will find the theory in the article on “Quantitative Genetics” in Wikipedia.
The distribution you have has a variance (a measure of the mean square of the deviation from its mean). For simple models of a quantitative trait whose value is a sum of contributions from many loci (genes) the variance, which is also called the phenotypic variance, is the sum of three components, the additive genetic variance, the dominance variance, and the environmental variance.
Each generation, each offspring is assumed to be the result of mating two parents, each randomly sampled from the population. This assumption is fundamental to quantitative genetics, it is the starting point for all the simple cases.
Let’s pause there for you to assimilate some of the terminology.
I appreciate it that you are modelling the reproduction of a population, rather than having us deal only with verbal descriptions. Let’s see if we can come to a meeting of minds.
In the above comment by me, the quotes by nonlin.org and me were mis-edited. The quote from “It’s still not clear’ on is by nonlin.org. I failed to catch this until after my ability to edit my comment expiref.
Nonlin.org,
Its Jock little pop-quiz time. That way he can say nothing and then pretend he is teaching you something, like he is the self-appointed expert on anything he says he is.
Hey Jock, what is the fittest creature on the planet? Just asking, see if you can answer. I will give you some multiple choice if you like. Take your time. Also please write a small essay to explain your answer.
This will be 20 % of your grade.
We all understand perfectly well. It is you that fails to understand the counterargument: Equally simple models exist that display regression to the mean, remain stable in the absence of perturbations, but will not return to the original population mean after an episode of directional selection. That runs counter to your argument that populations “regress to the old mean when the adaptive stressor is removed”.
Then, the question becomes which model is the more realistic. I think it is mine, because I have simply recycled basic quantitative genetic theory, which has for decades been succesfully applied in animal and crop breeding.
Yep, you definitely need to append smileys at the end of your humorous comments.
Joe Felsenstein,
Fixed it.
DNA_Jock,
Jock, you haven’t turned in your quiz results yet.
Need an extension?
For the guy that can’t read and/or understand:
1. Does the model already includes noise? Y/N?
2. What did I specifically wanted to remove from the model and why?
3. Did I manage to actually remove that item? Y/N?
4. Do you agree with “if it is true that (if A then B), and it is true that (not B), then it is true that (not A)”. Y/N?
5. Do you agree with “if divergence then spread, and if no spread then no divergence”. Y/N?
6. Do we see any spread of the distribution in nature over any observable time frame? Y/N?
7. Have I proved “regression prevents divergence” by turning off regression and seeing divergence”? Y/N?
Here you mangled the 7 statements above. In addition, you sneaked in claim A) for the first time in 381 comments and without proof. Good job.
You got all you need to duplicate the model and edit at will to prove your view point. For the 10th time: don’t ask me to do your job.
I appreciate that model. However, as explained, that model has a different purpose than mine, hence mine is as simple as needed to show what I wanted to show. We’re not inseminating cows for $ here.
Not my job to fix their model, but this assumption is BAD. How many organisms mate randomly? Humans certainly don’t do it. Doesn’t this conflict with “sexual selection”? Hey, it’s like the democrats debates: retards (retard concepts) fighting each other.
Not the first time he plays that little game.
Aren’t you forgetting something? Something very important and explained MANY, MANY times?
No doubt your model “runs counter”. But observations Trump your model. And observations show long time regression. Check them out… again:
Joke or no joke aside, you’re hereby forbidden to base your arguments on “fitness” until you prove your “fitness function”… unless you wear a clown hat.
nonlin,
My concern is that if I edit your model to demonstrate that you are hopelessly wrong, you won’t believe me.
If you were able to perform the edit I described, you would see the behavior I described [well, you will fail to understand the “regression to the mean” bit, but that’s just you. It will be there.] so we might be able to move past the “is too! is not! is so to!” and explore the limitations of your model together.
Instead, you will decline to engage, and just continue with your vague equivocations.
But in the spirit of continuing to “say nothing”, here goes:
I added noise with an sd of 0.25, just to make the regression to the mean more obvious.
Here’s the regression to the mean, and simultaneously the divergence of character.
The graphs
DNA_Jock,
I suspect it would be the kiss of death for me to say this seems eminently reasonable, so I won’t. 😉
Your model obtains the phenotype of the offspring by a mysterious process involving taking a single parent as somehow equivalent to two parents. It is not adequate to show what you wanted to show as the process of coming up with an offspring phenotype is mysterious.
Oh so now you want total realism? Actually you don’t have anything like that in your model, just a declaration that your reproduction by cloning from one parent is good enough.
Not interested in further discussion unless you want to clarify why your model works.
It always was, it seemed to me, a “bumblebees can’t fly” approach.
Except that, in the case of regression to the mean, the simple models show the bumblebee to be flying nicely.
No they don’t.
That is just false. None have been observed to “regress to the old mean” in the sense you mean. Please demonstrate this for any of those examples above.
For clarity: what you need to show to support your claim is a consistent bias in the mean offspring phenotype. You are not allowed to invoke differential survival and reproduction*. Renaming it “Design” won’t do either.
* unless you wear a clown hat, I understand.
Meaning that many species are currently not in their original form? That’s interesting. So how can you tell? For example, are horses currently in their original or domesticated form? Are they perhaps trying to regress to cute little eohippuses?
Why do you think this?
Because nobody in the history of genetics has ever reported this phenomenon, as far as I am aware.
Character reversals do happen, but are most parsimoniously explained by reversal of selection pressure. Antibiotic resistance is an even more complex story. I doubt that it is an appropriate example of character reversal, because there are many reports of microbial strains retaining their resistance upon return to a drug-free environment.
You’re not answering my questions and comments, but instead go on and on with your irrelevant monologue.
Sure it is very adequate and not at all mysterious. Let’s take height: In your opinion, what would be the average expected adult height of a M/F pair of siblings – given a population mean of 1.8 m – if parents are:
a. 1.8/1.8 m?
b. 1.8/1.4 m?
c. 1.5/1.5 m?
d. 2.0/2.0 m?
If you answer something like…
a. 1.8
b. 1.6
c. 1.5
d. 2.0
…then my model is PERFECT given I muted regression. Otherwise explain.
Not what I said. Read again.
Alan is stuck/stung by bumblebees. What’s the relevance to this analysis? That’s right. None whatsoever.
Sure they do.
Darwin’s finches cycle through beak sizes depending on the environment – they never go outside of the parameters observed already.
The peppered moth was white, turned black and then white again. Why not pink blue, or polka dot? This despite other moths having all kind of other colors.
Antibiotic resistant bacteria survives only in hospitals where it prays on the weakest of the weak. And only as long as stupid humans kill all other competing bacteria with antibiotics.
Domesticated plants & animals are helpless without CONSTANT human intervention. Feral animals never look anything like the pure breeds but instead more or less like their wild ancestors.
But you knew all this. Yet continue to refer to some crazy-ass “natural selection” and “fitness” that you’re not even able to articulate.
You have never seen any LIVE eohippus. So you have absolutely no proof that these are the ancestors of horses. Your circular reasoning fails.
Not only “do happen”, but MUST happen. If you disagree, give counterexample. Can you find any? Any at all?
Your “explanation” is BS and useless. Astrology is about “explaining”. Science is about PREDICTING.
Darwinist urban legend. If true, we would see antibiotic resistance everywhere, not just in the hospitals and other such weak spots. We don’t, hence your claim is FALSE. In addition, antibiotic resistance has ALWAYS been part of the biologic arsenal. And it will always be, so it will come and go. ALWAYS. Your “explanation” is useless.
So where do horses come from? And when?
If true, you’d be wrong? Merely that. Your extrapolations of what that would mean become invalid at that point, as you are already wrong on the basic point.
Okay nonlin, if you insist, I will answer your questions.
You might consider returning the favor…
No. 100% of variation is heritable in your model.
“regression to the mean”, because you misunderstand what it is.
Not really.
I do. Given how much time I spent teaching you precisely this point, it would be sad if I did not…
Yes, iff ‘divergence’ and ‘spread’ represent unambiguous and invariant true/false statements. (They don’t…)
Yes.
Given that your model can demonstrate (via the realistic addition of a little noise) BOTH ‘regression to the mean’ and simultaneously ‘divergence of character’, it is safe to say that you have proved that regression does NOT prevent divergence.
What you actually need is the swamping argument. You are over 100 years late to the party, however.
ETA to clarify: I took 5 to be a question about the validity of modus tollens, and not the truth value of any statements therein.
Nonlin.org,
This rant makes me wonder if Nonlin is being completely objective in his assessment of evolutionary theory. 😉
My first comment in this thread:
non-lin appears to be attempting a reincarnation of Fleeming Jenkin. Things have moved on a bit since the 1860s, non-lin.