
As shown repeatedly, “Nothing in Evolution Makes Sense. Period.” Not natural selection, gradualism, human evolution, UCD, tree of life, etc. And just to confirm, let’s look at another one of the nonsensical concepts of “evolution”.
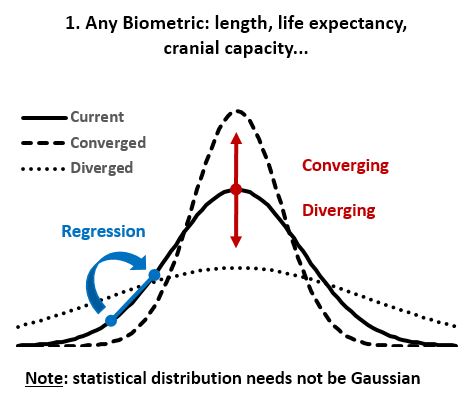

- ‘Divergence of character’ (character displacement or sympatric speciation) postulates: “during the incessant struggle of all species to increase in numbers, the more diversified these descendants become, the better will be their chance of succeeding in the battle of life. Thus the small differences distinguishing varieties of the same species, will steadily tend to increase till they come to equal the greater differences between species of the same genus, or even of distinct genera” (Darwin 1859). Sympatric speciation is hypothesized as “the evolution of a new species from a surviving ancestral species while both continue to inhabit the same geographic region”.
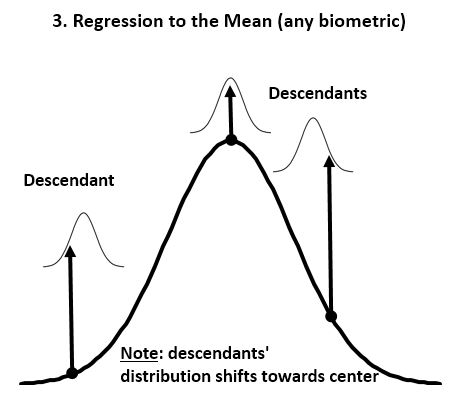
- ‘Regression to the mean’ is the biological law that overrules passive ‘Divergence of Character’. Any homogeneous population can be sorted statistically on various biologic metrics, usually resulting in a Gaussian (normal) distribution that is conserved over time in the absence of major environmental changes (as Mendel first showed; Fig 1&2). ‘Regression to the mean’ is thus the rule that causes the progeny of extreme individuals to be less extreme than their parents. Two outstanding tall parents will have statistically shorter children, and the progeny of the most and least intelligent/strong/aggressive/attractive/etc. will be more average than the parent. Many of the extremes have no descendants at all due to their limitations, and thus their “contribution” to the next generation is simply the average individual.
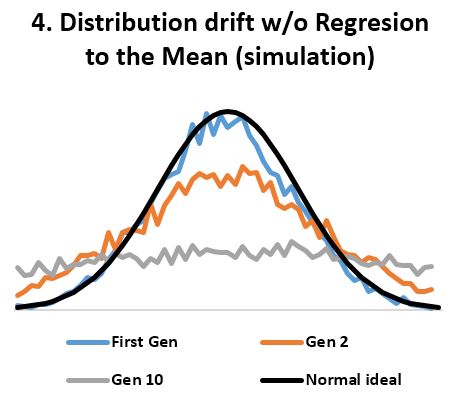
- In stable environments, population variability is extremely well conserved from generation to generation (Fig 3) as documented by the fossil and many other records. ‘Regression to the mean’ is thus a mathematical necessity without which a passive ‘divergence of character’ would be observed in very few generations (Fig 4). ‘Regression to the mean’ mechanism is incredibly accurate and allows for conservation of traits over thousands upon thousands of generations as observed. Scientists were rightfully surprised that ancient bacteria and many other fossils as well as mummified organisms including cats and monkeys are indistinguishable from their contemporary descendants. At a minimum, the number of organisms that show remarkable stability over long periods (living fossils) invalidate the ‘General Divergence’ theory. Does a limited, ‘Special Divergence’ hypothesis still make sense?
- Observed long term regression is highly unexpected and contrary to ‘divergence of character’ and ‘drift’ hypotheses. ‘Regression to the mean’ operates in the longest term observed, whenever environmental conditions are restored following significant changes that led to adaptive mutations. Most – if not all – organisms are endowed with a limited ‘plasticity’ trait that allows them to retain adaptive characteristics for generations. And yet, when the stimulus that caused the adaptation disappears, these organisms regress rather than maintaining those adaptive traits or accumulating even more diverging ones. Darwin’s finches, the peppered moth, antibiotic resistant bacteria and the domesticated plants & animals – all these and more have been observed to regress to the old mean when the adaptive stressor is removed, thus disproving even the limited, ‘Special Divergence’ hypothesis. These are not coincidences! The regression can happen over a few generations as in most epigenetic changes, many generations, and even the indefinite future if the adaptive stimulus is maintained (such as in domestication). Biologic variability can be compared to a loaded spring – the more it stretches, the harder the pull back (regression to the mean) and the more fragile is the extreme variant population. Domesticated plants and animals show that crossbreeds are resilient, while pure breeds are fragile showing that extinction of the extremes is the default outcome that promotes the ‘regression to the mean’ of the extended population.
- Adaptation neither demands not implies divergence in any way. What about the ‘adaptive radiation’ seen in Darwin’s finches, the cichlids of the African Great Lakes, and others? Is this not ‘divergence of character’? No. The driving force in all these and more is adaptation, not divergence even if “evolution” were true. Organisms just seek survival and, if their built-in yet limited plasticity matches the environmental challenges, these populations survive as variants. Otherwise, they simply go extinct like many others before. The new traits are not ‘divergent’ as shown by all known cases of reversals (as discussed) and none of further divergence when the adaptive stressor is removed. If ‘divergence of character’ were true, adaptive plasticity traits would be cumulative and sticky even after the adaptive stressor was removed, and the more extreme variants would be at least as resilient as the mean. Furthermore, experiments would show increasing variability over time in all research organisms and even more so in the short lived ones like bacteria. There would not be any distinct “species” and organisms would freely undergo metamorphosis (transmutation) into one another. Differential survival and randomness would eliminate all but the “best adapted” allele, therefore the Mendelian conservation of alleles would not be observed. Yet none of these are happening, thus falsifying the ‘divergence of character’ hypothesis.
- Adaptation is “fast and done”, “do or die” by necessity, unlike the supposed “slow and ongoing” ‘divergence of character’. If adaptation is not fast enough, the population simply goes extinct as many others did. The cichlids of Lake Victoria had less than 15,000 years to adapt and are as diverse if not more so than the cichlids in the other, much older African Great Lakes. But they do not need even that much time as the newer aquarium varieties obtained in a few generations show. Most likely, cichlids variants have come and gone throughout the history of all African Great Lakes in short cycles of adaptation. And that is why the cichlid biodiversity difference between a few years (Lake Victoria) and millions of years (other African Great Lakes) is unremarkable. The only remarkable fact is that cichlids have a predominantly Gondwanan distribution showing that in 180+ mil years, they did not adapt to ocean living despite their otherwise high adaptability. This clearly shows the limitations of adaptability and makes it an unlikely substitute to ‘divergence of character’. Darwin’s finches, peppered moths, bacteria, and many other also adapt fast or die as observed. And when the stimulus disappears, they revert just as quickly, and later readapt to whatever new stimulus they face or simply die out trying as confirmed. It is a very good thing ‘divergence of character’ is false, or else antibiotic resistant bacteria and other superbugs would have killed mankind by now as “evolution” falsely predicted.
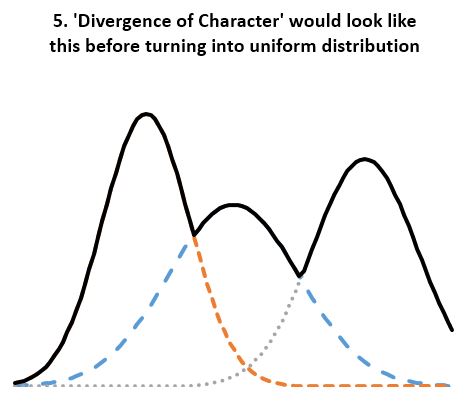
- Statistical evidence refutes ‘divergence of character’. According to the theory, “when organisms compete for scarce resources, natural selection should favor those individuals that are least like their competitors”. And since organisms always “compete for scarce resources”, the least average members of a homogenous population should always be favored by “natural selection”. If so, the well known normal distribution of any organism dimension (length, height, weight, etc.) should always be under pressure to change. We should see groups of “least like” the average form second, third, and so on normal distributions of their own, thus reshaping the original normal distribution into a composite distribution with several peaks and valleys as in Fig 5. And even that should not be adequate, as any concentration of similar individuals would be disadvantaged according to the ‘Divergence of character’ hypothesis, thus leading to uniform distributions as in Fig 4. However, neither Fig 5 nor uniform distributions are seen in homogeneous populations. Instead, we always see normal distributions. And since we see the normal distribution maintained over arbitrary number of generations and no hint of transitioning to a uniform distribution, the ‘Divergence of character’ hypothesis must be discarded. A trend not supported by several period observations must be discarded as noise artifact. This is the case for all examples considered including Darwin’s finches, the peppered moth, antibiotic resistant bacteria, cichlids, etc. All seem somewhat supportive of the divergence hypothesis over carefully chosen periods, yet the divergence is clearly illusory over longer periods.
- Are the bear of North America not like Fig 5? Yes, but they occupy different geographic regions. They are not homogenous. Indeed, we do encounter subfamilies of organisms, that have normally distributed metrics within the subgroup yet clearly distinct from those of other subgroups. However, where these subgroups overlap, the blend is always geographic and never biologic, meaning we see fewer of one kind and more of the other when moving from one’s territory to the others’ instead of blended characteristics as ‘divergence of character’ would predict. Humans are not different “species” although various subgroups are exclusively vegan/carnivorous, white/black, extra small/large. And domesticated organisms including canids are even more diverse than humans. Are the wild cichlids, finches, mice, and others qualitatively different than humans and canids? No. Then why the different “species”, many of which, ironically, are threatened by hybridization? The unwarranted inflation of “species” that do not even meet the loosest definition of reproductive isolation has the sole purpose of perpetuating the myth of ‘divergence of character’.
- Multimodal distributions in homogenous populations are not due to ‘divergence of character’. Indeed, bimodal distributions (Fig 2) and multimodal distributions are not uncommon in homogenous populations. However, these are due to the discreteness of physics in general and biology in particular, not due to ‘divergence of character’. Male and female populations are not diverging from one another and various alleles are in long term cyclical equilibrium as shown (spring model). ‘Drift’ is often invoked as a mechanism of ‘divergence of character’. This is wrong because ‘drift’ explains nothing as it is either aimless noise or due to adaptation and environmental change. Yet, as shown, adaptation is in no way ‘divergence of character’. In addition, the stable coexistence of several distinct variants within a homogenous population shows “gradualism”, “survival of the fittest”, and “natural selection” to be false because the alleles responsible are themselves distinct (no “gradualism”), they all “survive”, and neither is “selected” for or against.
- Darwin worried about regression to the mean for the wrong reasons. Namely, if blending inheritance (Darwin laid an egg) was true, then natural selection could not be true. Darwin puzzled over this a lot, but ended up with nothing satisfactory. Then Mendel showed that inheritance is discrete, not blended. Mendelian Inheritance Tables (see Punnett squares / Hardy-Weinberg equilibrium) show “probabilistic traits conservation” and thus disproving ‘divergence of character’ (at least as byproduct of reproduction) as well as dismissing “gradualism” (another one of Darwin’s unsupported claims).
- When entire populations split, do subgroups diverge from one another? This is not how ‘divergence of character’ is supposed to work.Descendants are supposed to diversify within the homogenous population. Furthermore, populations split by environmental conditions simply adapt to the new environment and for as long as those conditions allow. Adaptation is the driving force with no ‘divergence of character’ anywhere in sight. Island biology is the most diverse because islands are isolated and have many microenvironments. However, island variants are close descendants of their original colonists, showing that no divergence ever happened. Their risk of hybridization is high, disproving the “speciation” claim. They are also fragile examples of the extreme stretched biological spring model discussed, and will likely go extinct if at all stressed and when interacting with mainland.
Summary:
1. ‘Regression to the mean’ is the biological law that overrules passive ‘Divergence of Character’
2. In stable environments, population variability is extremely well conserved from generation to generation
3. Observed long term regression is highly unexpected and contrary to ‘divergence of character’ and ‘drift’ hypotheses
4. Adaptation neither demands not implies ‘divergence of character’ in any way
5. Adaptation is “fast and done”, “do or die” by necessity, unlike the supposed “slow and ongoing” ‘divergence of character’
6. Adaptation has limited powers and is thus not a substitute for ‘divergence of character’
7. ‘Divergence of character’ hypothesis would lead to uniform rather than normal (Gaussian) distributions as observed in homogenous populations
8. A trend not supported by several period observations must be discarded as noise artifact
9. The unwarranted inflation of “species” that do not even meet the loosest definition of reproductive isolation has the sole purpose of perpetuating the myth of ‘divergence of character’
10. Multimodal distributions in homogenous populations are not due to ‘divergence of character’
11. Mendelian tables show “probabilistic traits conservation”, disproving ‘divergence of character’ (at least as byproduct of reproduction), as well as dismissing ‘gradualism’
12. Island biology proves adaptation and the biologic spring model while disproving ‘divergence of character’
13. What’s in, what’s out? IN: ‘regression to the mean’, ‘adaptation’, coexisting variants, long term stability, spring model, normal distributions. OUT: ‘divergence of character’, gradualism, drift, speciation, uniform distributions, “natural selection”, “survival of the fittest”, “evolution”.
Links:
https://ucmp.berkeley.edu/bacteria/bacteriafr.html
https://www.sciencedaily.com/releases/2019/10/191018112136.htm
https://www.ncbi.nlm.nih.gov/pmc/articles/PMC3285564/
https://www.ncbi.nlm.nih.gov/pmc/articles/PMC3352989/
http://www.galton.org/essays/1880-1889/galton-1886-jaigi-regression-stature.pdf
https://en.wikipedia.org/wiki/Character_displacement
https://en.wikipedia.org/wiki/Sympatric_speciation
https://www.bionity.com/en/encyclopedia/Character_displacement.html
https://biologydictionary.net/divergent-evolution/
https://en.wikipedia.org/wiki/Cichlid
https://biology.stackexchange.com/questions/41982/regression-to-the-mean-and-evolution
Here we see the nonlin in his native habitat: the mixing of a purported ‘claim’ with an alleged observation (“sympatric speciation continues to do”), and a failure to notice that his model produces the very behavior he is asserting does not occur. Observe the intricately constructed, yet quite incoherent, strawmen he builds…
</David Attenborough>
This is utterly incoherent. You’re either totally clueless or you know you lack any good, logical arguments. My money is on “clueless”.
So where are we on the topic?
Kudos to Corneel and Allan Miller, the only two attempting some challenging questions, feeble as they were. However, it looks like they both gave up – unlikely due to them seeing the light – but rather for regrouping in search of better arguments. The worse would be if they learned nothing. Sadly, this is also a very strong possibility.
Refers to this:
and your killer rejoinder:
“Logical”, you keep using that word…
You must be ready to take on the World, now. 😉
Let’s see:
To your 1) How about “a hypothesized phenomenon that has not been demonstrated”?
To your 2) How about “a theory proposed by Darwin and rebranded Sympatric Speciation”, hence not needing a new definition?
To your 3) True, but with what purpose? Do you REALLY think I built a model to disprove myself? What would be the chance of that? Hmm…? Could it be something else? Say:
If ‘divergence of character’ were TRUE, then normal distributions would disappear into uniform distributions within x generations (x = 10 in this model). But since normal distributions DO NOT disappear (they conserve), if follows that ‘divergence of character’ is FALSE.
I DO keep using that word… Am I wrong to do so?
You continue to use “divergence of character” to refer to a phenomenon and to a theory, proving my point about your confused thinking. Although equating “divergence of character” with “a theory proposed by Darwin and rebranded Sympatric Speciation” represents a new pinnacle. Yikes.
It seems you did not realize that YOUR simulation shows normal distributions with ever widening variances. News flash: your simulation (and your figure 4) does not show any ‘uniform’ distribution.
When, in the OP, you state
you are completely and utterly wrong.
Evidently so.
Just for clarification, how does “Divergence of Character” relate to “Character Displacement“?
ETA: Never mind, I see the OP has as first line
confirms it is the same concept – sympatry.
Just thinking “out loud”:
An experiment to test sympatric speciation. The hypothesis to test is if you take a genetically homogeneous population of organisms, split it into representative groups and keep them separately in identical environments and observe changes over time; evolutionary theory suggests that mutations and other variations in genomes would arise and new alleles would spread if beneficial, lost if lethal or deleterious, and neutral variation may be fixed or lost by genetic drift and the isolated populations would become different from each other. You’d need something small with fast generation times but it could be done.
Oh wait…
Richard Lenski beat me to it!
But I can see this doesn’t test sympatric speciation, just the general case of breeding isolation. Might it be that examples of sympatry are really peripatry at the microniche level. Someone mentioned upthread the apple maggot as an example of sympatry but one could suggest apple fruit are a separate niche from hawthorn berries. It’s a bit like the distinction between natural, artificial and sexual selection. It’s really the same process at the level of alleles.
Opportunity to specialize to separate ecological niches is indeed a relevant aspect. But the other problem to consider, as always, is sex. Without assortative mating, subpopulations are incapable of adapting to their respective niches, since recombination will continuously break apart adaptive allele combinations (Joe may have some thoughts on this topic). Only if there is some mechanism that reduces the exchange of genetic information, then linkage disequilibium is allowed to build up. In the case of the apple maggot fly this is accomplished by assortative mating on the host fruit. If you feel this is a bit cheating, then let me tell you there are other possibilities, such as sexual selection. In such cases females of one subpopulation discriminate against males of the other. The beauty of this scenario is that mate discrimination is adaptive, and will be selected for, since matings with the “wrong” partner result in unfit offspring that cannot thrive in either niche.
ETA: clarity
Word to the wise: I haven’t really addressed Nonlin’s comments about sympatric speciation, because it’s a red herring. Despite being a fascinating topic, we should be aware that sympatric speciation has rather specific requirements, and is therefore uncommon. The most common reason for divergence of subpopulations remains geographic isolation.
(attributed to Theodosius Dobzhansky)
No, it seems perfectly reasonable. I was just suggesting, from the maggot fly’s perspective, the apple fruit and hawthorne fruit are separate niches. Allopatric speciation rather than sympatric (thougn the quibble seems semantic and the distinction is somewhat arbitrary).
There was another example given of speciation in sticklebacks where one incipient species adopted bottom feeding and another mid-water which makes the allopatry/sympatry distinction similarly arbitrary (depends on how tightly you construct the niche boundaries. I take the point about sexual selection, too.
I agree and that causes me to wonder why Nonlin wants to focus on the exception rather than the rule. Isolated populations will diverge genetically, even if the environment is stable.
In some ways, I am surprised that there isn’t more sympatric speciation, given the various forms of assortative mating that we observe.
Explanation 1: these species are, in fact, heading for a speciation event, and we should alert R. Byers (and his descendants) to monitor the situation over the next thousand years.
Explanation 2: there is counterbalancing disassortative mating effect that becomes more marked as the sub-populations diverge. MHC, anyone? You need to maintain a positive feedback effect for the sub-species to go all the way.
The burning question: how will nonlin incorporate these effects in his model?
DNA_Jock,
Even if there is not disassortative mating, if there are two populations with the genotypes having different fitnesses, the gene frequencies in them will diverge some. However that does not necessarily continue if there is gene flow between the populations. They can come to an equilibrium state where changes due to selection are balanced by changes due to gene flow. This can occur even with assortative mating. Any imperfect assortment, when there is also gene flow, will result in an equilibrium being reached.
In 1981, a mere 39 years ago, I wrote a paper on such a model, following up on models by others such as John Maynard Smith (1966), It can be found here (I hope the link works for everyone).
Joe,
Thank you for the link – I have only had time to scan it, but I have downloaded it for further perusal. Very cool.
Whilst educating myself about your Santa Rosaria reference, I came across this slide deck.
Slide 28 might amuse you…
ETA: I guess I was envisioning cases where the associative mating is extreme, and stays that way…
Well, I did deliberately play off of the title of his famous paper, so yes, it is descended from that paper in that sense. I apologized to him in the Acknowledgements for the abuse of his paper’s title. I also sent him a preprint, again apologetically. He didn’t express any dissatisfaction with it.
I had heard him give a paper at a symposium in 1969, on how one ought to define species in asexual organisms. Very nice work. He really was one of the greatest ecologists.
What “phenomenon”? I demonstrate there’s no such thing. Only a failed theory.
It only simulates 10 gen and it approximates very well the uniform distribution. And anyone can see where this leads. Combine that with natural hard limits on both ends (fact. and as intentionally presented in fig 4) and the uniform distribution is inevitable. Your legalistic argument fails.
Btw, if your argument had any strength, you would not have waited for 250 other comments. But kudos for the effort. You did wake up apparently.
Not how this particular hypothesis is supposed to work. Probably another topic.
As explained, nothing arises, nothing is beneficial or deleterious, and there is no genetic drift. Why would you bother commenting on this topic if you ignore everything discussed? And there’s no natural or sexual selection. The idea of sexual selection in particular is totally retard.
Are you trying to sneak in a concession? Why so late?
I do discuss geographic isolation too and why that is not “divergence” but adaptation – see 8. and 11.
Are you even thinking about addressing these:
4. Adaptation neither demands not implies ‘divergence of character’ in any way
5. Adaptation is “fast and done”, “do or die” by necessity, unlike the supposed “slow and ongoing” ‘divergence of character’
6. Adaptation has limited powers and is thus not a substitute for ‘divergence of character’
?
I explained. Perhaps you missed:
“Furthermore, populations split by environmental conditions simply adapt to the new environment and for as long as those conditions allow. Adaptation is the driving force with no ‘divergence of character’ anywhere in sight. Island biology is the most diverse because islands are isolated and have many microenvironments. However, island variants are close descendants of their original colonists, showing that no divergence ever happened. Their risk of hybridization is high, disproving the “speciation” claim. They are also fragile examples of the extreme stretched biological spring model discussed, and will likely go extinct if at all stressed and when interacting with mainland.”
I am addressing a particular topic. Plus you don’t read for comprehension.
Nope. “The unwarranted inflation of “species” that do not even meet the loosest definition of reproductive isolation has the sole purpose of perpetuating the myth of ‘divergence of character’.”
Way ahead of you. That “counterbalancing” force that prevents sympatric speciation is called Regression to the Mean.
You keep BS-ing about “fitness”, but will never tell us what YOUR “fitness” is.
A real life example would give a modicum of credibility to you otherwise fantastic and non-scientific scenario.
Awkward.
[Remember, nonlin, this is using your mind-blowingly poor model…]
The concept of the fitness of a genotype (or a phenotype) in an environment will be meaningful to most readers here. If nonlin.org does not think that there is such a thing, that’s sad, but I have tried before to talk sense into nonlin.org, to no avail. Maybe someone else can persuade nonlin.org that it is sensible to talk in terms of fitness.
If fitness does not differ from one genotype to another, or from one phenotype to another, then all of evolutionary biology will be a wasted effort. Hundreds of thousands of scientific papers gone up in smoke, Departments closing, scientists wandering the streets unemployed. Course curricula needing to be changed massively.
Or else possibly nonlin.org is wrong about fitness.
Joe Felsenstein,
This appears to be the abyss evolutionary biology is staring into. Why are we identifying proteins of animal groups (mammals) with no mutations becoming fixed in the population.
Alpha actin 1.
Human Mouse Rat alignment 100%
The abyss into which I am staring is one that is supposed to contain an explanation of colewd’s argument. I see little in there, all right. Yup, that is one quite conserved protein. Histones have other, similar examples.
But how does one make that into an argument against evolution? I can only guess, but my best guess is that colewd concludes from this that no other sequence could possibly be functional enough to allow evolution to get closer and closer to this conserved sequence. But that does not follow at all from the strong conservation. It’s very echoey in there …
colewd,
Like saying you’ve disproved the notion of travel by observing that some people have never been anywhere. Between conserved proteins and orfan genes, one wonders if there is anything left for which we might even require a theory of change … 🤔
According to a certain Bill Cole, that is because any sequence change will be causing system failure, i.e. purifying selection against variants that confer low fitnes.
I have told you this several times, but let’s repeat it once more: Your appeal to purifying selection as the reason for sequence conservation entails accepting both fitness variation and the efficacy of natural selection.
colewd,
By the way, Nonlin thinks that your claim that Design involves the generation of de novo sequences is complete bollocks:
I trust the two of you will politely discuss this, and inform us of your consensus afterwards.
Oh, for crying out loud. Why do you need to argue against every single point that is raised? Jock is simply right. The process in your simulation will never ever result in a uniform distribution. Don’t believe me? Test it yourself! Simulate as many generations as your computer can handle and look at the resulting distribution! It will be a Guassian!
Jock is right, you are wrong. Accept the correction, say “thank you” and move on. Sjeesj!
Not so fast. Actually you missed the qualification nonlin.org made:
So it is apparently assumed that there are “hard limits” at the edges of the graph beyond which the phenotype does not go. If those are reflecting barriers (rather than absorbing barriers) the normal distribution’s tails will fold there. The distribution, as it spreads, will flatten out, and ultimately approach a uniform distribution.
Dna_jock should try this in the simulation that made the graph that Dna_jock most recently showed. Any individual that exits the interval [-8, 8] gets bounced back in from those values by the amount it moved past the boundary.
Oh I did already.
Well I didn’t have the boundary reflect, rather I modeled ‘Stop at the Wall’: a bounded random walk.
And the result is much as you describe, Joe.
I was waiting for nonlin to argue that his “natural hard limits” were required for his model to perform as he described it.
You have revealed my carefully-laid trap. Mind you, given nonlin’s difficulties with map/territory, he may fall into it yet…
Scuse questions from someone rather math-challenged but:
Am I following correctly? Nonlin’s spreadsheet, as clarified by DNA_Jock, demonstrates exactly the opposite of what he is claiming? Without feedback, just a little randomizing at each generation eventually produces a flat line between the limits that we might call a “divergence of character”?
How quaint!
Yes Alan, and (insofar as I can understand nonlin) his point appears to be that we NEVER SEE THIS “divergence” in the real world. Due to a powerful force called “regression to the mean”, which defeats Darwinism. Apparently.
Those naughty evolving populations are going to stay where their Creator intended them to be!
So, Nonlin’s mission, should he choose to accept it, is to show us how “regression to the mean” works, what the driving force is – maybe with a spreadsheet model.
Nonlin? Do we get to include inheritance, variation or selection yet?
Allan Miller,
The data base is continuing to reveal conserved proteins over millions of years of “evolution”. Why are they so conserved? Is it mass purifying selection or is it the fact the amino acid substitutions that could work require too many steps for the mutations to get fixed in the population?
Evolution cannot work without lots of variation in order to explore solutions.
And? There’s obviously just the right amount as here we are along with the rest of living diversity (though threatened by accelerating climate change and habitat destruction) on this planet.
Alan Fox,
How did this produce alpha actin 1?
PS @ Bill
If you are going to respond along the lines of “no, not evolution but the product of a mind” maybe you can start a new thread and include answers to the conundrum of why God needed to create a Universe when he only needed the Sun and this planet, why he needed to create all life if only humans are ensouled and why he didn’t just pop us all straight to heaven and save all the bother. Is the answer “God moves in mysterious ways”?
Do you mean “how did alpha actin evolve”, Bill? I don’t know. I don’t know whether there is a proposed pathway by successive small steps favoured by selection. Can you tell me it is impossible because of some specific barrier to evolution in this case. Or is it the usual “too complex to have evolved” rule that you apply as a general copout.
Alan Fox,
Then I ask how did the spliceosome evolve or the ubiquitin system. Your answer is again I don’t know. If I ask how did light from a star deflect while traveling near the sun the scientific theory of general relativity can give me an answer.
On what is the basis of your belief that it did evolve? In order to evolve it first needs to get fixed in a population and this step does not appear to be trivial. When most amino acid mutations appear to be more often deleterious from a population perspective.
It’s revealing far more variation. As usual, you stare at one thing and ignore the rest.
The most highly conserved tend to be structural (eg actin, histone, ribosomal) or informatic (eg ubiquitin). There’s far more variation in (most) enzymes.
There are two principal reasons for conservation, one is being pinned in place by other interactors, restricting latitude, the other being they’re already as ‘good’ as they can get.
Yes, but if it doesn’t find any paths in a given clade for a given protein, is that a problem? It has the rest of the genome to tinker with.
The argument you are advancing, clearly fallacious, is that evolution could not have happened in the past for these proteins because you can’t change them now.
colewd,
Questions are easier to ask than answer. (To the relief of many, I’m sure) I have no more time to respond today. Perhaps tomorrow.
Allan Miller,
This is not the argument. You are invoking a straw-man and this is what I was testing. If you have to do this you have no legitimate theory. The design hypothesis is necessary to stop the evolutionary stories that have no grounding in empirical evidence.
The problem I am surfacing is the challenge of getting mutations fixed in populations is not trivial especially in highly functioning species like. vertebrates. How does a fish population evolve the ability to breathe.
Someone at PS produced this Bill Cole Bingo card for times like now when Bill starts blithering his usual ID-Creationist talking points.
Sadly it doesn’t take long to fill up at all.
It is precisely the argument. You and gpuccio have repeatedly used the current/recent conservation of proteins as evidence of their unevolvability. It is utterly bogus, for reasons that have been explained to you many times.
Well, that’s quite the non-sequitur you are rocking, mate. He could be really bad at arguing. He could be arguing in his spare time.
Strange, then that 99.9% of the people who generate the empirical evidence are backing the evolutionary stories over the design hypothesis.
What is “the challenge of getting mutations fixed”? Please be very specific, as it appears to me that you don’t understand what fixation means.
Adapa,
ROFL. Priceless!
No, not a strawman. It may not be your intended argument, but if you see conservation as a problem for evolution, that argument is the one you are making.
You need a theory with no empirical evidence to deal with a theory with (according to you) no empirical evidence?
Conserved sequences are irrelevant to this – unless you are making the argument you just said was a strawman.
How does a Gish develop the ability to gallop?
Just shows the shakiness of evolutionary theory that one man could destroy it by uttering a logical fallacy! 🤣
Fish breathe via numerous mechanisms which one are you confused about?