
As shown repeatedly, “Nothing in Evolution Makes Sense. Period.” Not natural selection, gradualism, human evolution, UCD, tree of life, etc. And just to confirm, let’s look at another one of the nonsensical concepts of “evolution”.

- ‘Divergence of character’ (character displacement or sympatric speciation) postulates: “during the incessant struggle of all species to increase in numbers, the more diversified these descendants become, the better will be their chance of succeeding in the battle of life. Thus the small differences distinguishing varieties of the same species, will steadily tend to increase till they come to equal the greater differences between species of the same genus, or even of distinct genera” (Darwin 1859). Sympatric speciation is hypothesized as “the evolution of a new species from a surviving ancestral species while both continue to inhabit the same geographic region”.
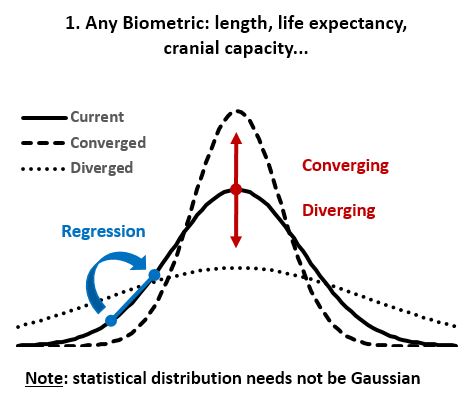
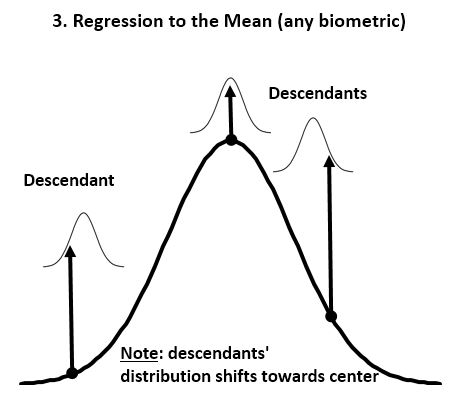
- ‘Regression to the mean’ is the biological law that overrules passive ‘Divergence of Character’. Any homogeneous population can be sorted statistically on various biologic metrics, usually resulting in a Gaussian (normal) distribution that is conserved over time in the absence of major environmental changes (as Mendel first showed; Fig 1&2). ‘Regression to the mean’ is thus the rule that causes the progeny of extreme individuals to be less extreme than their parents. Two outstanding tall parents will have statistically shorter children, and the progeny of the most and least intelligent/strong/aggressive/attractive/etc. will be more average than the parent. Many of the extremes have no descendants at all due to their limitations, and thus their “contribution” to the next generation is simply the average individual.
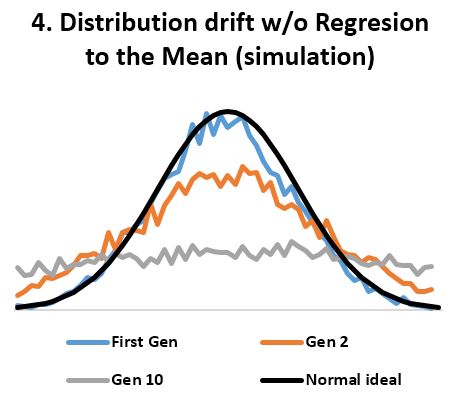
- In stable environments, population variability is extremely well conserved from generation to generation (Fig 3) as documented by the fossil and many other records. ‘Regression to the mean’ is thus a mathematical necessity without which a passive ‘divergence of character’ would be observed in very few generations (Fig 4). ‘Regression to the mean’ mechanism is incredibly accurate and allows for conservation of traits over thousands upon thousands of generations as observed. Scientists were rightfully surprised that ancient bacteria and many other fossils as well as mummified organisms including cats and monkeys are indistinguishable from their contemporary descendants. At a minimum, the number of organisms that show remarkable stability over long periods (living fossils) invalidate the ‘General Divergence’ theory. Does a limited, ‘Special Divergence’ hypothesis still make sense?
- Observed long term regression is highly unexpected and contrary to ‘divergence of character’ and ‘drift’ hypotheses. ‘Regression to the mean’ operates in the longest term observed, whenever environmental conditions are restored following significant changes that led to adaptive mutations. Most – if not all – organisms are endowed with a limited ‘plasticity’ trait that allows them to retain adaptive characteristics for generations. And yet, when the stimulus that caused the adaptation disappears, these organisms regress rather than maintaining those adaptive traits or accumulating even more diverging ones. Darwin’s finches, the peppered moth, antibiotic resistant bacteria and the domesticated plants & animals – all these and more have been observed to regress to the old mean when the adaptive stressor is removed, thus disproving even the limited, ‘Special Divergence’ hypothesis. These are not coincidences! The regression can happen over a few generations as in most epigenetic changes, many generations, and even the indefinite future if the adaptive stimulus is maintained (such as in domestication). Biologic variability can be compared to a loaded spring – the more it stretches, the harder the pull back (regression to the mean) and the more fragile is the extreme variant population. Domesticated plants and animals show that crossbreeds are resilient, while pure breeds are fragile showing that extinction of the extremes is the default outcome that promotes the ‘regression to the mean’ of the extended population.
- Adaptation neither demands not implies divergence in any way. What about the ‘adaptive radiation’ seen in Darwin’s finches, the cichlids of the African Great Lakes, and others? Is this not ‘divergence of character’? No. The driving force in all these and more is adaptation, not divergence even if “evolution” were true. Organisms just seek survival and, if their built-in yet limited plasticity matches the environmental challenges, these populations survive as variants. Otherwise, they simply go extinct like many others before. The new traits are not ‘divergent’ as shown by all known cases of reversals (as discussed) and none of further divergence when the adaptive stressor is removed. If ‘divergence of character’ were true, adaptive plasticity traits would be cumulative and sticky even after the adaptive stressor was removed, and the more extreme variants would be at least as resilient as the mean. Furthermore, experiments would show increasing variability over time in all research organisms and even more so in the short lived ones like bacteria. There would not be any distinct “species” and organisms would freely undergo metamorphosis (transmutation) into one another. Differential survival and randomness would eliminate all but the “best adapted” allele, therefore the Mendelian conservation of alleles would not be observed. Yet none of these are happening, thus falsifying the ‘divergence of character’ hypothesis.
- Adaptation is “fast and done”, “do or die” by necessity, unlike the supposed “slow and ongoing” ‘divergence of character’. If adaptation is not fast enough, the population simply goes extinct as many others did. The cichlids of Lake Victoria had less than 15,000 years to adapt and are as diverse if not more so than the cichlids in the other, much older African Great Lakes. But they do not need even that much time as the newer aquarium varieties obtained in a few generations show. Most likely, cichlids variants have come and gone throughout the history of all African Great Lakes in short cycles of adaptation. And that is why the cichlid biodiversity difference between a few years (Lake Victoria) and millions of years (other African Great Lakes) is unremarkable. The only remarkable fact is that cichlids have a predominantly Gondwanan distribution showing that in 180+ mil years, they did not adapt to ocean living despite their otherwise high adaptability. This clearly shows the limitations of adaptability and makes it an unlikely substitute to ‘divergence of character’. Darwin’s finches, peppered moths, bacteria, and many other also adapt fast or die as observed. And when the stimulus disappears, they revert just as quickly, and later readapt to whatever new stimulus they face or simply die out trying as confirmed. It is a very good thing ‘divergence of character’ is false, or else antibiotic resistant bacteria and other superbugs would have killed mankind by now as “evolution” falsely predicted.
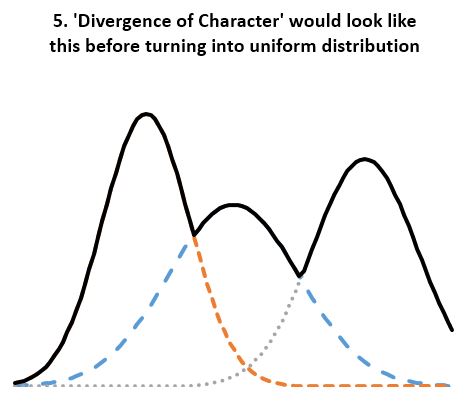
- Statistical evidence refutes ‘divergence of character’. According to the theory, “when organisms compete for scarce resources, natural selection should favor those individuals that are least like their competitors”. And since organisms always “compete for scarce resources”, the least average members of a homogenous population should always be favored by “natural selection”. If so, the well known normal distribution of any organism dimension (length, height, weight, etc.) should always be under pressure to change. We should see groups of “least like” the average form second, third, and so on normal distributions of their own, thus reshaping the original normal distribution into a composite distribution with several peaks and valleys as in Fig 5. And even that should not be adequate, as any concentration of similar individuals would be disadvantaged according to the ‘Divergence of character’ hypothesis, thus leading to uniform distributions as in Fig 4. However, neither Fig 5 nor uniform distributions are seen in homogeneous populations. Instead, we always see normal distributions. And since we see the normal distribution maintained over arbitrary number of generations and no hint of transitioning to a uniform distribution, the ‘Divergence of character’ hypothesis must be discarded. A trend not supported by several period observations must be discarded as noise artifact. This is the case for all examples considered including Darwin’s finches, the peppered moth, antibiotic resistant bacteria, cichlids, etc. All seem somewhat supportive of the divergence hypothesis over carefully chosen periods, yet the divergence is clearly illusory over longer periods.
- Are the bear of North America not like Fig 5? Yes, but they occupy different geographic regions. They are not homogenous. Indeed, we do encounter subfamilies of organisms, that have normally distributed metrics within the subgroup yet clearly distinct from those of other subgroups. However, where these subgroups overlap, the blend is always geographic and never biologic, meaning we see fewer of one kind and more of the other when moving from one’s territory to the others’ instead of blended characteristics as ‘divergence of character’ would predict. Humans are not different “species” although various subgroups are exclusively vegan/carnivorous, white/black, extra small/large. And domesticated organisms including canids are even more diverse than humans. Are the wild cichlids, finches, mice, and others qualitatively different than humans and canids? No. Then why the different “species”, many of which, ironically, are threatened by hybridization? The unwarranted inflation of “species” that do not even meet the loosest definition of reproductive isolation has the sole purpose of perpetuating the myth of ‘divergence of character’.
- Multimodal distributions in homogenous populations are not due to ‘divergence of character’. Indeed, bimodal distributions (Fig 2) and multimodal distributions are not uncommon in homogenous populations. However, these are due to the discreteness of physics in general and biology in particular, not due to ‘divergence of character’. Male and female populations are not diverging from one another and various alleles are in long term cyclical equilibrium as shown (spring model). ‘Drift’ is often invoked as a mechanism of ‘divergence of character’. This is wrong because ‘drift’ explains nothing as it is either aimless noise or due to adaptation and environmental change. Yet, as shown, adaptation is in no way ‘divergence of character’. In addition, the stable coexistence of several distinct variants within a homogenous population shows “gradualism”, “survival of the fittest”, and “natural selection” to be false because the alleles responsible are themselves distinct (no “gradualism”), they all “survive”, and neither is “selected” for or against.
- Darwin worried about regression to the mean for the wrong reasons. Namely, if blending inheritance (Darwin laid an egg) was true, then natural selection could not be true. Darwin puzzled over this a lot, but ended up with nothing satisfactory. Then Mendel showed that inheritance is discrete, not blended. Mendelian Inheritance Tables (see Punnett squares / Hardy-Weinberg equilibrium) show “probabilistic traits conservation” and thus disproving ‘divergence of character’ (at least as byproduct of reproduction) as well as dismissing “gradualism” (another one of Darwin’s unsupported claims).
- When entire populations split, do subgroups diverge from one another? This is not how ‘divergence of character’ is supposed to work.Descendants are supposed to diversify within the homogenous population. Furthermore, populations split by environmental conditions simply adapt to the new environment and for as long as those conditions allow. Adaptation is the driving force with no ‘divergence of character’ anywhere in sight. Island biology is the most diverse because islands are isolated and have many microenvironments. However, island variants are close descendants of their original colonists, showing that no divergence ever happened. Their risk of hybridization is high, disproving the “speciation” claim. They are also fragile examples of the extreme stretched biological spring model discussed, and will likely go extinct if at all stressed and when interacting with mainland.
Summary:
1. ‘Regression to the mean’ is the biological law that overrules passive ‘Divergence of Character’
2. In stable environments, population variability is extremely well conserved from generation to generation
3. Observed long term regression is highly unexpected and contrary to ‘divergence of character’ and ‘drift’ hypotheses
4. Adaptation neither demands not implies ‘divergence of character’ in any way
5. Adaptation is “fast and done”, “do or die” by necessity, unlike the supposed “slow and ongoing” ‘divergence of character’
6. Adaptation has limited powers and is thus not a substitute for ‘divergence of character’
7. ‘Divergence of character’ hypothesis would lead to uniform rather than normal (Gaussian) distributions as observed in homogenous populations
8. A trend not supported by several period observations must be discarded as noise artifact
9. The unwarranted inflation of “species” that do not even meet the loosest definition of reproductive isolation has the sole purpose of perpetuating the myth of ‘divergence of character’
10. Multimodal distributions in homogenous populations are not due to ‘divergence of character’
11. Mendelian tables show “probabilistic traits conservation”, disproving ‘divergence of character’ (at least as byproduct of reproduction), as well as dismissing ‘gradualism’
12. Island biology proves adaptation and the biologic spring model while disproving ‘divergence of character’
13. What’s in, what’s out? IN: ‘regression to the mean’, ‘adaptation’, coexisting variants, long term stability, spring model, normal distributions. OUT: ‘divergence of character’, gradualism, drift, speciation, uniform distributions, “natural selection”, “survival of the fittest”, “evolution”.
Links:
https://ucmp.berkeley.edu/bacteria/bacteriafr.html
https://www.sciencedaily.com/releases/2019/10/191018112136.htm
https://www.ncbi.nlm.nih.gov/pmc/articles/PMC3285564/
https://www.ncbi.nlm.nih.gov/pmc/articles/PMC3352989/
http://www.galton.org/essays/1880-1889/galton-1886-jaigi-regression-stature.pdf
https://en.wikipedia.org/wiki/Character_displacement
https://en.wikipedia.org/wiki/Sympatric_speciation
https://www.bionity.com/en/encyclopedia/Character_displacement.html
https://biologydictionary.net/divergent-evolution/
https://en.wikipedia.org/wiki/Cichlid
https://biology.stackexchange.com/questions/41982/regression-to-the-mean-and-evolution
Would you do me a favor, please? Whenever Bill Cole shows up here in the comments again, please repeat this exact phrase to him.
Thanks
Disingenuous! Plant and animal breeders have selected preferred specimens of their crops and flocks and bred from them. It is all the same thing, natural and artificial selection. If farmers design then the environment designs – and the environment designed farmers.
Which is utter bullshit. This has been discussed extensively and it is not the topic of the day. Go comment there if you want. But read first: http://theskepticalzone.com/wp/natural-selection-evolution-magic/
Can’t be. A “first” is alone by definition. Quit speculating about the distant past. You’re ridiculous.
Which is and was an utterly retard idea as discussed. Go read.
You did not and I’m encouraging laziness: the genetic bottleneck natural experiments are good approximations of the Adam and Eve mouse. Point is, the genetic material recovered nicely over time.
You’re making up stuff again. As long as the finches remain finches there’s no “evolution”. And don’t give me the crap about your inner fish and inner worm and inner cockroach and whatnot.
You read but didn’t understand. Ex: you can’t take a mean of blue, brown, green, etc eyes. Can you take the average of those with and without the BRCA gene? Apply the Mendelian tables to these variants and see they’re in static equilibrium. “What about mutations?” you ask. Well, do those mutations collectively amount to anything or just noise? And here we’re talking about “divergence or character”, so same environment.
God (is my belief).
A long time ago. That long.
More wimpy equivocation bullshit. Cut it out. Make it algebra, not bull-crap.
But nobody is saying it was “a first” except you, my clueless friend
Two aspects, not two types is my observation. Share if you know more/different.
What “evolutionary change”? Senseless nonsense of the third grade.
It just does. Experimentally. If you like it or not. “Why” is the wrong question at this point.
What are you smoking? You’re miss-interpreting and miss-applying. I’ve had enough repeating: “reality TRUMPS your models”.
1. Keeping genotype constant beyond 10 gen.
2. It’s simplistic for something you clearly don’t understand.
3. You abuse it to extract more than it can do given 1. and 2.
…
…
4. As opposed to mine: a simple model => an indisputable VERY LIMITED conclusion.
5. I didn’t model regression (pretending to know something I don’t like you). I modeled a world WITHOUT regression and showed that to be contrary to reality.
Won’t remember. You do it. What’s his point?
How the heck (to not say fuck) is this “disingenuous”?
The farmers design because they have a goal in mind. They want more eggs, cheese, meat, etc. They want finer wool, faster growth, etc.
Does your “environment” have a goal in mind? I thought not.
A retard that knew nothing of the topic once said “nature is a tinkerer not a designer” or some like that. Other retards copied the nonsense not knowing tinkerers are designers and designers are tinkerers. Anyone who has ever built anything at all in his/her life should know this basic fact.
Then don’t talk about “biological Eve”. Too late complaining once you buy into the concept.
No, you’re ridiculous! 😛 Mitochondrial Eve is not proposed to be the first human female, nor is Y chromosome Adam the first human male. You really need to grapple with some basic genetics.
“Go read, go read”. You are turning into something of an automaton.
A definition is simply that. It doesn’t have to describe something true. If something is evolution as commonly defined, then sticking your stupid scare quotes round it and whining “no it’s not” isn’t getting you anywhere.
Which genetic bottleneck experiments are those?
Call it what you like, if adaptive mutations can arise from a bottlenecked/founder population, you are describing the very thing you are railing against.
Righto … 🤔
Indeed you can’t. So how do we determine whether a population statistic for such a trait is the perturbed or the ‘regressed’ mean? If the former, how does the latter establish itself?
Ahem … “quit speculating about the distant past. You’re ridiculous”. 🤣
“Mitochondrial” Eve, not biological Eve
Yes, she is: http://www.macroevolution.net/mtdna-human-chimpanzee.html
Same.
Words have meanings in the real world. Apparently not in the “evolutionary” fake world. Because equivocating is convenient. This BS won’t fly.
This time you have to go read. Go search old comments.
There’s a very important distinction between adaptation and divergence even if you don’t like it:
Perturbed by what? Mendel showed that the ratio of alleles is not changing if the environment is constant per this case (divergence…) discussed here.
I was asked for my opinion and clearly stated so. Unless you clearly do the same, expect push-back on your BS.
Are you really that dense? (rhetorical question)
Nothing in that article suggests that Mitochondrial Eve was the only existing woman of her time, let alone the first one to ever exist.
Because you’re struggling, here are a few more considerations about my model and why it works while yours doesn’t:
1. I model a normal distribution for something like height (nothing fancy). Is it realistic? Yes. Check.
1B. Do I need to know how regression works? No. Check
2. The population is constant. Not growing, not decreasing. One average parent gives birth to only one average child. Is it realistic? Yes. Check.
3. The distribution of the progeny is skewed to the center. Do I need to model? No because I am doing away with that for the model. Check.
4. Is the distribution of the progeny same as the initial distribution of the whole population? No, it is much narrower. Does it matter? It slows down the decay of the normal into uniform from 10 gen to whatever depending on the parameters. Will the final result be different? NO. Check.
5. Does the result invalidate a general “divergence of character”? Yes. If ‘divergence’ were true, the distribution would flatten as per the model. The distribution does not flatten, therefore ‘divergence’ is not true.
6. Did I miss anything? No. Check.
The passage I quote does. True, it’s a guy citing another guy and disagreeing with him. But I don’t care being an agnostic about their “evolution” debate. Fact is, if the Mitochondrial Eve hypothesis still stands (and I believe it does), then by necessity, we have one original woman which is what Eve stands for.
No, it doesn’t.
Nope. “Mitochondrial Eve” as a name was invented by the San Francisco Chronicle science writer John Tierney in 1986. He had heard that Allan Wilson was going to give a research report talk to his department, the Biochemistry Department at University of California, Berkeley. He attended this. Allan Wilson and his group had done trees of mitochondrial restriction-site data on 147 individuals of various ethnicities. They showed that the root of the tree, the female-line ancestor of all 147 samples, was in Africa about 200,000 years ago.
To be provocative, Tierney wrote this up for the next day’s paper with the provocative headline “Scientists Find Eve”. I know. I happened to be driving to San Francisco the morning that appeared and I read that headline while eating breakfast in Yreka, California. Later that day I got to Berkeley and called Allan’s lab, as I always liked to check in with them about what new results they had. Allan’s student Mark Stoneking answered the phone, and when I asked for Allan, said “Allan’s in hiding”.
Now if you know any genetics, you know that the genes in the rest of the genome, all the nuclear genes, come from different ancestors than just the female line. So there is no requirement that they have the same common ancestors (most recent common ancestors of their copies) as that “Eve”.
But “knowing any genetics” may be too much to ask.
I’m sure most everyone reading here realizes this is just another example of nonliin making his/her ignorance known to the world.
Without even delving into the published literature on the subject a look at the wiki page for Mt eve and Y-adam provides insight on how wrong nonlinear is:
https://en.wikipedia.org/wiki/Mitochondrial_Eve
and
https://en.wikipedia.org/wiki/Y-chromosomal_Adam
oops…JoeF beat me to it!
Always interesting to hear stories like this!
My memory is faulty. I checked. The story in the San Francisco Chronicle was by Charles Petit, not John Tierney. It appeared on March 24, 1986 under the title “The Mother of us All — A Scientist’s Story” and used the phrase “African Eve”.
No she isn’t. It literally says it in your quote:
“They think that all modern mtDNA can be traced back to a single female in the ancient population”
Unless that population otherwise consisted of only males, then she wasn’t the ‘first female’. And did none of those males have mothers?
Yep. And in the real world outside of Creationism, no-one defines evolution as you do. So the equivocator is you.
Can’t be arsed. If you can’t support your claims, you’re bluffing.
Yes, there is. Regardless, when you talk about cichlids, you’re talking about them using existing variation to diversify. But you’re also saying that that variation can arise by mutation. Ergo …
Perturbed by whatever force you imagine stretches your ‘spring’. Or does it only work on quantitative traits?
I know, just having a bit of fun at your expense. You can’t act like a dick and expect others not to do the same.
So, god did it and it happened a long time ago.
6000 years ago is a long time. Is that a sufficiently long time?
My argument is you are attempting to differentiate one process into two by equivocating with the word “design”. Plant and animal breeders do not design; they select. If you want to call plant and animal breeding design then I am equally entitled to say the environment designs. It’s the same process – a change in allele frequency in populations.
When (or if) people are able to design and produce DNA sequences that they can insert into genomes and correctly predict the outcome of such insertions, then I might agree there is real design going on. Work on GMO is still currently at the stage of test-it-and-see.
Evolution is a process resulting from a change in allele frequency brought about by differential reproduction influenced by the niche environment. That’s my understanding of the consensus view on biological evolution. Your attempts to claim evolutionary models fail as explanations are not persuasive to me, or to anyone else as far as I can see. Recall I asked you a while ago whether anyone was at all taken with your output on this issue. You replied “Yes” but offered no link or other evidence. What was your justification for answering “Yes”?
Yes. Selection.
Rumraket,
Indeed!
To complete my comments on the original work on Mitochondrial Eve, the scientific paper on this was by Rebecca Cann (now of the University of Hawai’i), Mark Stoneking (now of the Max Planck Institute for Evolutionary Anthropology in Leipzig), and the late Allan Wilson. Reference is:
Rebecca L. Cann, Mark Stoneking & Allan C. Wilson. 1987. Mitochondrial DNA and human evolution. Nature 325: 31–36.
It’s behind a paywall, but here is a 2010 interview with Becky Cann about the origins of the project and reaction to it.
(And, by the way, the Max Planck Institute where Mark Stoneking is located was for a long time headed by another Allan Wilson student, Svante Pääbo, and is often abbreviated as MPI-EVA.)
So you’re just saying the “Mitochondrial Eve” hypothesis doesn’t stand. Fine. I couldn’t care less, even though – contrary to your claim – I keep hearing about this hypothesis.
Anywho, this is a tangent on a tangent totally unrelated to the ‘divergence of character topic. Now can we get back to what matters?
I’m glad you see the truth.
What could this mean: “existing variation to diversify”? I am not saying “variation can arise by mutation”. Variation doesn’t “arise” like hocus-pocus. It’s an inherent (built in) capability of all organisms.
Well, we got regression to the mean that accounts for the spring model. But that doesn’t apply to alleles as Mendel showed. If you think there’s a MEASURABLE and DIRECTIONAL “perturbation” in allele composition, it’s your task to describe and measure this phenomenon. Then we’ll think about explaining it.
Note: I’ll ignore the other nonsense you keep repeating. Maybe you get the message.
Completely irrelevant trivia. On the other hand, it’s VERY surprising that you, that claim so much knowledge, have nothing to say about the ‘divergence of character’ myth. How come?
No, it’s not the “same process”. Look again – what did I say the big differentiator is when it comes to design? You can dispute, but at least make sure you understand what you read.
You have never designed a thing in your life? Be honest!
You can change alleles all you want and will never have “evolution” as Darwin dreamed. Your understanding is bad. Very bad.
Dude, we’re talking about ‘divergence of character’ here. Do you even understand how this is supposed to work?!? You’re obviously clueless about the topic. Hint: ‘sympatric’ versus ‘allopatric’ “speciation”.
One more thing: what are you trying to demonstrate? Be clear and concise! I asked this before and still didn’t get a clear answer. Add this to the list of failures of your model.
No, a number of us here were pointing out that statements other people make about Mitochondrial Eve did not mean what you thought they meant.
OK, so we’re agreed that once again, you were talking about something you didn’t understand. Because you immediately wanted to stop talking about it.
I’ll add it to *some* list of failures allright. You seem to be forgetting that “my” model is a minor modification of your model. The only thing I introduced was a clear separation of heritable and non-heritable variation. Doing so, one can easily demonstrate that regression to the mean is a consequence of the non-heritable componenent of variation, whereas the response to selection is accomplished by a change in heritable variation.
This is an incredibly obvious and intuitive result. High school students get it, first year biology students get it, heck at some point even you got it:
Precisely! And since the change is heritable, it will not revert, in contrasts to your insistent and completely counter-factual claim that it will. You even invented some magical “second aspect of regression to the mean” that should do the job. I pose that this second aspect of regression to the mean does not exist and is solely the fruit of your imagination. The only things that will reverse a genetic change are the mechanisms of evolutionary change: natural selection, genetic drift, mutation and selection.
I’ll address the specific objections to the modified model here. Note that I have renumbered the second part for ease of reference.
There is no “beyond generation 10” in your model, so there was no need to model it.
Parsimony is considered a virtue, and a sign of elegance.
Guilty as charged, but not really a valid objection, is it?
Sure, but a valid conclusion nonetheless. You still haven’t conceded the point.
Agreed, your original model lacked realism, but after I modified the model it showed regression to the mean, exactly as in empirical findings.
Nonlin: [H]ere are a few more considerations about my model and why it works while yours doesn’t:
Agreed.
False. Please educate yourself on variance, covariance, correlation and regression.
False. Real populations have unequal family sizes, and hence experience genetic drift. I agree it was sensible to not include drift in the model though.
This is an imaginary property on your behalf. The progeny distribution within families will be normal as well, but centered on the parental genotypic value, not its phenotype.
Haha, the distribution of the progeny within families is identical to the initial distribution of the whole population. You are sampling from the same distribution. Please revisit point 6B.
False. The flattening in your model is a result of the population constantly generating new heritable variation. This is a non-realistic assumption. I have told you this before. Please pay attention to me when I tell you smart things.
I’ll give you a hint: It starts with “S” and rhymes with “election”.
Amusingly, you are both wrong. Eye colour is a quantitative trait, not a strictly Mendelian one. The colour of your eye is largely determined by the amount of pigment produced by the melanocytes in your irises, a decidedly continuous trait. The appearance of a Mendelian pattern of inheritance is created by the large impact of segregation of allelic variation at the OCA2 gene, a major locus for eye colour.
But you can score eye colour on a blue-green-brown scale, which I did at one time during my classes. In this crude experiment, the trait had a heritability of roughly two/third of the phenotypic variation.
The moral of this story: you are both wrong to claim that Mendelian traits cannot be analyzed as quantitative traits. The only requirement is that the trait can be quantified on some continuous scale.
You have variation arising in your ‘mouse Adam and Eve’s descendants. It’s not inbuilt; mA & mE didn’t have it and their descendants did. The capacity to generate it might be, but there is no direct evidence that mutations anticipate need.
Mendel showed no such thing. He didn’t apply his work to populations. In populations, allele frequencies can change, clearly and measurably. Mendel was looking at isolated breeding lines, without population processes.
The message that you’re not competent to discuss this subject? Yep, got that loud and clear.
Corneel,
I didn’t claim that Mendelian traits can never be analysed quantitatively. I started off talking of hypothetical A’s, B’s and C’s: genotypes, necessarily discrete character states in themselves, regardless of their phenotype. Nonlin brought up eye colour as a f’rexample; delving into the genetics of eye colour would have been (and is) a distraction from the point I was pursuing.
“it was a tree with a high likelihood of being correct and it was consistent with a lot of other data —anatomically modern fossil forms in South Africa around 200,000 years ago.
There is something very familiar about South Africa…
I think I’ve read it somewhere, in an old book perhaps…
John Harshman would probably know where exactly a certain garden was located…
Can you imagine what would happen, if the old book said the mitochondrial Eve came from Israel? Or Germany?
All this work is proving what had been known all along…
One of very few examples of not wasted research money…
It’s easy enough to sample a population and estimate allele frequencies. That’s the MEASURABLE part. If one samples a later population, it doesn’t have to be DIRECTIONAL, it just has to be ‘different’ – not what it was. And the question becomes, which of those statistics is the ‘right’ one – the one to which the population would return if further distortion occurred? If you want a concrete example, the Drosophila P element is as good as any. What’s the frequency of P to which left-alone populations would ‘regress’?
Fair enough. He certainly failed to grasp the concept in his “Discrete versus Gradualism” thread.
Yes it is!
Selection, natural, artificial and sexual, is all the same process.
Same process. Speciation happens when populations cease to share genes, which can happen by geographical isolation or behavioural isolation.
You’re just scrambling to undo a big oops. Very transparent.
“Once again”? Do you have a list?
It’s a far off tangent that I don’t care about, but since you are so arrogant, let’s note that no one disproved the hypothesis of ONE AND ONLY ONE EVE. Let’s also note all claims on this are motivated by the religious belief of the claimant be him/her atheist, evolutionist or creationist. Hence must be taken with a lot of salt. Also let’s observe that the human mitochondria is of ONE type, not two (as is the human sperm). This clearly indicates a single origin.
The bigger mystery remains “why is Joe silent on divergence?” “Did he not understand the topic”? “Does he think divergence is not salvageable?”
I strongly disagree.
We discussed. How would you know if that’s realistic? Prove it.
Also there is NO SELECTION. “Divergence of character” assumes homogeneous populations in constant environment. Enough with the nonsense!
Only you don’t get it, Corneel. So ironic.
1. See above – there’s no chop off in “divergence”.
2. We see this long term regression all the time in all the examples given and more. If you like it or not. Can you come up with an example where long term regression is not happening?
3. “mechanisms of evolutionary change” is not the answer even in theory. In at least one of the examples given, there should not be a regression to the same precise anterior design, but an “evolution” away in one of the myriad other directions. This aside from “mechanisms of evolutionary change” being totally bogus.
We got a stand-off because you’re in denial. What now?
The failure to communicate appears to be yours.
Try harder to support some of your assertions? Provide some indication that someone, anyone, is taking your claims seriously? Find another hobby?
You’re doing something different, so your requirements are different. Btw, what the heck are you doing? Because if you just assume the design (genotype) is constant for ever, you’re just proving my point that there’s no divergence as we’re all unchanging living fossils. Is this what you want?
Only if it makes sense. Else it’s dumb like yours. When you go on chopping off the distribution, your stunted distribution lives for ever which is known to be false experimentally.
“Forced conclusions” is not “a valid objection”?!?
Not my model. The model of those that proposed “divergence”. Just because you curve fitted to the same output doesn’t mean your assumptions are realistic.
Why false? Where’s your proof that’s “false”?
“Drift” it’s just another word for “noise” unless you prove otherwise. You can’t say “false” and then agreeing with my approach. This is not about the “family size”, but about the population size – which is constant in long term constant environments.
Any proof? If not, BS.
This is dumb. If both parents are tall, the progeny is far more likely to be tall than a midget. Only not more extreme. This is known experimentally.
How so “false”? This is what “divergence of character” proposes. Of course “divergence of character” is not realistic as shown. Because we don’t see that outcome, it’s true, not false that the result invalidates a general “divergence of character”. Pay attention to the goal.
Same bad comment Alan Fox made. Why are you not reading?!? Btw, explain to Alan Fox why he continues to be clueless. If you get it, that is.
Claims that are not supported with evidence can be dismissed without evidence.
A model that doesn’t include selection is not going to be of much use in arguing that evolutionary theory – in which biased sampling of alleles is core to outcomes – is incorrect.
There’s no selection in your model. There is selection in reality and selection is – together with variation – a pillar of evolutionary theory.
When you’re right, you’re right. Frame it cause it isn’t often.
Anyway, you should agree with the other example: a regression to the mean of w/out BRCA gene doesn’t make sense.
Don’t push your luck. I didn’t say “Mendelian traits cannot be analyzed as quantitative traits”. Read again.
Of course it is (the capacity). “Anticipate need” is not what we were discussion.
Mendel showed probabilities – which you can only get from populations.
The issue is not “allele frequencies can change”, but whether that change is more than noise in a constant environment (as required when considering “divergence”).
Return when you complete your task:
“If you think there’s a MEASURABLE and DIRECTIONAL “perturbation” in allele composition, it’s your task to describe and measure this phenomenon.”
What are you smoking? Another topic you abandoned. That’s why you’re still clueless.
I have nothing new to add anymore, so I’ll bow out of the conversation at this point. You should just go on pondering the significance of genetic variation and heritability. If you don’t learn to see the relevance of genetics in evolutionary theory, you are doomed to be forever attacking strawmen.
It is a very strange case study.
It looks like nonlin cannot distinguish between a set of observations, a proposed explanation, and a model that (accurately or inaccurately) attempts to represent certain aspects of reality.
His lack of understanding of basic statistics and basic biology leads him to create in his mind strawmen that are so outlandish that no-one can recognize the argument that he is attempting to disparage.
He certainly fails to understand what evidence is, and how models might be validated…
By way of example, he appears to be using the phrase “divergence of character” to describe
1) a natural phenomenon (whose existence he denies)
2) an evolutionary theory that predicts this phenomenon (under circumstances undefined by nonlin)
3) the behavior of his only disclosed simulation
OTOH he uses “regression to the mean” to refer to
1) a natural phenomenon (which he incorrectly claims is universal)
2) an undocumented force of nature that causes this phenomenon (due to his failure to understand high school math)
3) something which he has claimed to have modeled — no idea where, though…
I reckon that vagueness of language (c.f. Sal) and map/territory is sufficient to explain the entirety of his confusion, but I might be missing something.
Here you go again. But quitting is indeed much better than going in circles.
Ridiculous to tell me to ponder when this whole essay is based on deep thoughts on variation and heritability.
Perhaps you should learn to dissociate the REAL science of genetics from the FAKE, pseudo-science of “evolution”.
On another note, for those that are still confused about “divergence of character” relation to “selection”, this is Darwin being even more retard than his usual self. What he claimed – and sympatric speciation continues to do – is that a homogeneous population will continuously split and “diverge” even when the environment doesn’t change, hence no “selection pressure” is applied on said population. In this blunder, Darwin burned his escape route – as flimsy as it is – to “natural selection”.
You are too badly confused to discuss this. When you wake up, go back, read, and try to understand what is said, where the evidence lies, and what the logical conclusions are.
Ask not where the evidence lies
But why it lies to thee.