
We keep hearing that Darwin is obsolete and “evolution” has turned into a real science lately. Apparently not. Here is why this relatively new theory – that was supposed to rescue “evolution” – is in fact demonstrably incorrect.
- Drift in biology is “evolution by randomness”. The ‘drift’ idea was championed by Motoo Kimura as “The Neutral Theory of Molecular Evolution”. Kimura was motivated by JBS Haldane’s dilemma: too much polymorphism exists for “natural selection” to be able to pick and chose “beneficial” genes due to gene colocation in the genome and to the propagation time in the population. Of course, Haldane’s dilemma is only due to the conflict between his nonnegotiable assumptions that “natural selection”, “fitness” and “beneficial mutations” are all real and the observable realities of biology. The ‘drift’ idea claims that random events cause allele frequencies to change over time and in extreme cases, ‘drift’ is seen eliminating or fixing certain alleles in the whole population. Drift is supposed to operate independent of “natural selection” to the extent the allele is not strongly “beneficial” or “detrimental” for its frequency to be primarily a product of “natural selection”.
- Outside of “evolution”, drift adds to nothing or is aimlessly carried by known forces. In the first case it has no effect on anything either in the short or the long term. In the second case it is explained by the carrying force and therefore is itself not an explanation of anything. Examples of driven drifts are driftwood, wind-driven snow, rain, cloud, dust, or smoke as well as aircraft moved by air currents (all these driven). Inconsequential drift examples include aimless course or random movement. In biology too, genetic drift should either be inconsequential or, if lasting changes are observed, not drift but the real driving forces behind this drift should be held responsible.
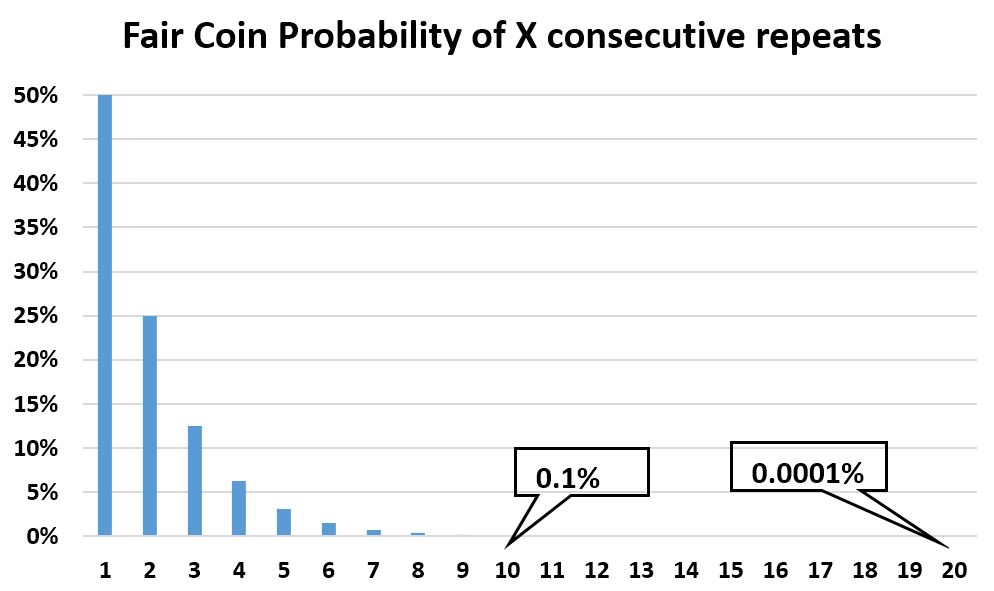
- A fair coin is a good model for inconsequential biologic drift. But how can that be, given mutations can take the population in a myriad directions, not just two (head and tails)? The fair coin model works because from a starting population, there are only two mutually exclusive directions possible on any of the myriad dimensions: towards or away from the initial value. For instance, mutations can make a second generation taller, greener, heavier, etc. But the third generation can only be even more tall, green, heavy, etc. than the original population, or less so than the second generation and therefore more like the original population. Just as with a fair coin, the output is binary: further away (call it ‘tails’) or closer to (call it ‘heads’). To prove random drift irrelevant in biology, let us consider the statistical case for drift by using a fair coin sequence. Yes, there is a statistical probability however small to see a series however large of tails (or heads), but eventually that series must end. As the chart shows, the probability of a sustained random drift decreases exponentially to nil. And over a much longer time, the realized events still sum up to an average of “no change”. This is easily verifiable experimentally. And if a significant bias persists over the long term, then the coin cannot possibly be ‘fair’.
- Population bottleneck cannot lead to “evolution”. Populations sometimes decrease dramatically due to competition, disease, predation or natural catastrophes. But killing the majority of one allele while sparing most of another is either a non random scenario or not possible except in the tiniest of populations and thus inconsequential. Post bottleneck, mutations occur and are claimed to further cause the strain to diverge from the original. But if no driving force is involved, there is no reason to believe mutations would work in a diverging direction only as the statistics discussion proves. Therefore, only a driving force, not drift can be responsible for long term changes following a bottleneck event. Alleles could indeed disappear from a population due to a series of random events, but either the population is small hence the loss is irrelevant; or, if that small population grows, a mutation will likely reintroduce the allele; or, in an already large population, the allele loss is highly unlikely due to random events not scaling up with the population size. Either way, random allele loss cannot possibly drive “evolution”.
- Founder effect also cannot lead to “evolution”. Many islands contain unique species due to having been populated by only a few organisms. Similar to bottlenecks, founder effect reduces the amount of genetic diversity in a population, meaning certain biological characteristics are overrepresented and these individuals are hypothesized to form new populations with different gene pools from the original population. But the same problem arises: the sum of mutations cannot be divergent unless a force other than drift itself causes that. If the environment weren’t drastically different, rather than diverge one would expect the new population to converge to the old strain instead given that at least that strain is known to be viable. Furthermore, if one small group found its way to the island, what prevents other small groups from repeating the trip and cancelling any such founder effect? The best explanation for the founder effect is thus that it is an ephemeral artifact and that all lasting differences are just adaptations unrelated to any random drift.
- There is more to the “dilemma” than Haldane’s take, and it’s all bad for “evolution”. A few things we know for sure: a) bottleneck populations are much more homogeneous as observed in the northern elephant seal, European bison, domestic animals, etc.; b) unique founder features (including injurious ones) persist for a long time in the founded population c) established populations (no bottleneck) are likely to be polymorphic with morphs in long term equilibrium. The theory of “evolution” predicts “beneficial” mutations taking over the population resulting in an “evolved”, better population over time. This is inconsistent with polymorphism and with the propagation of the founder’s features (some seeming “deleterious”) over time. Not only that, but normal population variability argues against “natural selection”. Because “fitness” functions cannot be calculated even in principle, let alone forecasted for any of the morphs, a hypothesized “natural selection” cannot possibly operate in biology. Furthermore, newfound populations, while adapted to their new environment, are in no way superior to their mainland cousins, even after the longest time from the split. This clearly shows “beneficial mutations” impossible. Thus, Haldane’s dilemma can easily be solved by removing his prejudiced, hopeless and disproved assumptions of “natural selection”, “fitness” and “beneficial mutations”.
- Balancing selection, an alternative to drift, fails to solve Haldane’s dilemma. The big idea of “balancing selection” is that the “fitness” of an organism with two alleles (heterozygote) is higher than the “fitness” of either allele (homozygote). Perhaps “fitness” changes in time from young to adult. Or “fitness” of a phenotype is dependent on its frequency relative to other phenotypes – rare morphs of prey are actually “fitter” due to predators concentrating on the more frequent morphs. However, there is no way to forecast anything of this sort or even to explain with any confidence the frequencies of these alleles. An elusive, fluid “fitness” attempting to explain all ends up explaining nothing, therefore it is a redundant hypothesis. And why would “fitness” changes in time? Why wouldn’t a morph “evolve” to do away with this temporal weakness? And what should the frequency and number of alleles be? Again, no rhyme or reason. Take ABO blood system. Why not more or fewer types? Why these frequencies? And why the current distribution? These are not just unanswered; they are unanswerable question with any degree of confidence.
- In conclusion, both drift and “natural selection” are disproven by polymorphism. As seen, drift in itself is not an explanation. All bottleneck drift “examples” are of theoretical cartoonish type (colored balls/ rabbits/ bugs). Some founder effect examples are real but inconsequential (northern elephant seal and Amish polydactyly) as clearly none of these leads to any “evolution”. Meanwhile, “natural selection” is perplexed and for sure inexistent as “fitness” is nowhere observable let alone quantifiable, therefore also a chimera.
Links:
https://en.wikipedia.org/wiki/Haldane%27s_dilemma
https://en.wikipedia.org/wiki/Neutral_theory_of_molecular_evolution
https://en.wikipedia.org/wiki/Genetic_divergence
https://en.wikipedia.org/wiki/Balancing_selection
https://en.wikipedia.org/wiki/Heterozygote_advantage
https://en.wikipedia.org/wiki/Polymorphism_(biology)
https://en.wikipedia.org/wiki/List_of_polymorphisms#Ants
https://en.wikipedia.org/wiki/Heterozygote_advantage#Cystic_fibrosis
https://sciencing.com/difference-between-natural-selection-descent-modification-17942.html
Not a good start, what can we expect but misunderstanding and misinformation from the rest?
I suppose “we” here means Nonlin, and “keep hearing” means Nonlin misunderstands something. Of course Darwin’s initial presentation of evolution and proposed mechanicms had to be incomplete, thus obsolete, that doesn’t mean that it wasn’t science. It was and continues to be science. That new discoveries improve prior ones, and/or fix problematic parts of older ones, doesn’t make the prior attempts any less scientific. You need a good education Nonlin. Start with reading for comprehension.
Of course not. It has been science from the very start.
Wow, you’re very misinformed Nonlin. Drift is not supposed to rescue evolution. Evolution doesn’t need rescuing. Drift is just supposed to help understand some facts that the prior set of described evolutionary-related phenomena didn’t cover. That’s it.
This is as misinformed as talking about Newton’s “law” of gravitation only becoming science once Einstein’s work “rescued” it, just because Newton’s theory failed to account for some of the facts it was trying to describe, and for lacking a mechanism that would explain how gravitation works at a distance. Newton’s take on gravitation was missing something, that didn’t mean that there was no gravitation, it only meant that as science progresses other elements can be added to the theory and mistaken parts fixed.
This misinformation is just the introductory paragraph. is it worth examining the rest? Am I going to find more misinformation? I bet Nonlin will not fail to disappoint in being obtuse, misinformed, and that she will be unable to process any corrections. After all, that’s been Nonlin’s MO each and every time.
Entropy,
#3 is my favorite.
I continue to be impressed at the various branches of science and mathematics that nonlin.org has refuted. Can publication and a trip to Stockholm be far behind?
DNA_Jock,
Well, given that Nonlin’s concept of mutation is one where a single mutations is instantly shared by a whole population, what else can we expect if not a trip to Stockholm?
ETA: Holy crap! I just read #3 in full, and, as wrong as it is, Nonlin cannot be consistent with her own model. She first explains that the fair coin would have to fall differently for the population to remain mostly unchanged, meaning that the result would be drift/neutral, only to finish declaring that drift doesn’t work because the coin doesn’t always fall on the same side!!!
ETA after ETA: Nonlin will not be able to understand how she’s contradicting herself.
Most of this bullshit of an OP can be summarized as an incoherent diatribe whose only purpose is to demonstrate that neutral theory is wrong because: it’s neutral!!!!!!
🤣😂😂🤣😂
nonlin.org quoth:
That’s quite remarkable. Now I had thought that when we keep track of the number of Heads while making independent tosses of a fair coin, there would be two possible outcomes: more Heads or fewer Heads. Not toward or away from the initial value. In modeling genetic drift in a population of finite size, say 1000 diploid individuals, so 2000 copies of the gene with two possible allels, we start out with some gene frequency of one allele, such as 0.30. Then the next generation is modeled as produced by 2000 tosses with the Heads probability 0.3. If the result is a fraction 0.33 of Heads, the gene frequency is now 0.33. Then for the next generation we toss 2000 times with Heads probability 0.33, not 0.3 (and certainly not 0.5). And so on. The resulting gene frequency of the allele that corresponds to Heads will wander up and down until it ends up at 0 or at 1. It does not tend to head back to 0.3 and stay there.
So nonlin.org has misunderstood how tossing coins relates to random genetic drift.
If nonlin.org gracefully acknowledges this and says “Yes, I guess I was wrong about that” I will be utterly flabbergasted and you can knock me over with a feather.
I’m understanding this to mean that 0 means nobody has it, and 1 means everyone has it. But does it have to “end up” at either one, or if it’s gone and reappears, is that consider a novel instance? If everyone has it and then some start to lose it, is that possible? In other words, are 0 and 1 truly terminal conditions?
In the case where there are no mutations or gene flow into the population, once it gets to a frequeny of 0 or 1 then the population stays there forever. That was the simple case I was discussing.
If the missing allele recurs due to mutation, then the population will continue wandering between 0 and 1 and the distribution of how often it is at various values can calculated from the forward and backwards mutation rates and the effective population size. As to whether it is then “considered a novel instance”, that is just a matter of what terminology you prefer.
GENETIC DRIFT!
If you are going to make a half-baked argument against a process, you should at least get the name right.
But shouldn’t the change of getting Heads be larger because of regression to the mean? After all, according to you populations always return to their initial value, right?
My intuition says that if heads get off by chance to a big head start (so to speak), the overall percentage of heads will decrease simply because a 59% chance is less than the current percentage. So over time, with enough tosses, two things happen: the total count difference between heads and tails will increase, and the percentage difference will decrease.
You’re not showing any connection. Do you lack conviction because you got burned so many times?
But is your model true to reality? Polymorphs say otherwise. And your 1000 individuals is a small population. Perhaps a Critically Endangered in the Extreme population. Furthermore, we can go all the way back to Mendel and see allele stability.
My point exactly in par. 4.
Alan Fox,
Calm down. It’s a failed theory anyway, so you won’t hear about it much longer 🙂
Not “always”. As I explained back then, just “whenever environmental conditions are restored following significant changes that led to adaptive mutations.” Or as I explain here, “But the same problem arises: the sum of mutations cannot be divergent unless a force other than drift itself causes that.”
And there are no guarantees in statistics.
Flint,
This is not clear. Neither ‘heads’ nor ‘tails’ is favored in a fair coin. With populations, if say starting height is 175, many ‘tails’ can take it to 180, while many ‘heads’ can take it to 170. Either one can be divergent, but if the coin is fair, then equilibrium is restored… eventually… at least in the model.
Yeah, I am disappointed but not surprised that you fail to see the connection. It’s a reminiscent of the “HGP helps drug discovery” story, where the connections were explicitly laid out by Entropy and myself, but nonlin failed to comprehend.
Here it is a little more straightforward: your OP is about modeling random walks. This is well understood math, and you get it profoundly wrong in a number of ways.
I will restrict myself to the hilarious para3.
Three errors :
A) You equate a walk in multiple dimensions with a walk in one dimension. This is wrong.
B) You equate a walk in one dimension with the probability of a consecutive series of steps in one direction. This is wrong.
C) You claim that the P(N consecutive steps in one direction) “decreases exponentially to nil.” as N increases. Nope, the to nil bit is obviously wrong, even for the geometric series you are displaying.
In fact, Flint’s ‘intuition’, viz:
is a well-known mathematical result. It is “experimentally verifiable”, too. 😀
Hence my link.
Just because nonlin.org cannot see the connection, does not mean that the connection does not exist, or has not been demonstrated. Your ignorance is evidence of your ignorance, nothing more.
Genetic drift is a well-documented fact.
Neutral theory proposes that most of the mutations that end up fixed in the population are, ahem, phenotypically neutral. So why would a “low probability” of a non-neutral scenario, namely “heads-after-heads-after-heads-after-heads”, prove neutral theory wrong?
——–
Polymorphisms say that mutations continue to happen and that fixation/elimination don’t happen instantaneously, not that no mutation ever gets fixed or eliminated from a population.
If you have problems with small populations, then, to be coherent, you’d have to renounce your own “coin” model, which implies a “population” of one per generation. Either that or your mutations are fixed as soon as they happen, in a single generation.
That either fixation or loss will eventually happen doesn’t mean that they happen in just a few generations. Look:
Again, reading for comprehension Nonlin. Very important.
I admit I didn’t come in at the beginning, but I’m not sure I understand. What does “phenotypically neutral” mean?
A while back, I visited a museum exhibit of medieval suits of armor. I was impressed with how small they were — today I thought any male over the age of 10 would be too large to fit. Similarly, literature of a few hundreds years back describes someone 6 feet tall as a giant of a man. The size increase has been attributed to several contributing factors, but it’s real.
So does this trend indicate a violation of neutral theory, or a refutation of regression to the mean, or does it mostly indicate better diet?
Yes, important to include the improvements in nutrition, which are also, btw, the main driver of earlier menarche. The guys-are-bigger-these-days effect is real, but is over stated by the suits of armour. Fun fact: children’s costume armour is more likely to survive intact to the present day than actual, adult working armour, which tends to get dinged up and recycled. There is therefore some ascertainment bias at work too.
It means that the organism’s characteristics are not affected much.
As DNA_Jock explained, most if not all of that seems explainable by changes in environment, like better nutrition, less severe pathogenesis, etc. Either way, neutral theory is about most mutations, not about every mutation. Some other Japanese, I think she was a student of Kimura’s, but not sure, proposed nearly-neutral theory, since there’s, clearly, a good possibility of mutations with slight effects that can survive for quite long and even get to fixation according to simulations in population genetics.
As I said, even if those changes you describe have, at least in part, a genetic basis, it would be covered by the rest of mutations. Those with phenotypic effects. Neutral theory is not absolutist. We leave those kinds of black/white ideas to the likes of Nonlin, who thinks, for example, that dictionaries rule reality, rather than humans putting definitions into dictionaries, not made precisely of stone, just to try and convey ideas to one another, and changing them, adapting them, as needed.
Your intuition is correct, but mind that with genetic drift there is no 50-50 chance of drawing one or the other allele; you have a higher chance of drawing the most common allele. The chance of drawing a given allele equals its current allele frequency, which changes every generation. To return to the initial frequency there needs to be some memory of what that initial value actually was. Of course, no such memory exists. Hence, Nonlin’s dichotomy of “towards or away from the initial value” is a red herring.
I believe that in your current OP you argue that populations always return to their initial value, because the statistical probability of “up” and “down” equals out in the long run. So how does that differ from “regression to the mean”?
Corneel,
@ Flint, you might like to try one of the simulations on offer.
The University of Washington, Seattle sim mentioned here or OM’s sim here, which is illustrative of how unpredictable the result is between the two last surviving alleles. In the sim I ran, the loss of alleles was rapid until the final two, frequency changed several times above and below 50% and fixation took much longer.
(Not sure how close to reality the model is)
Hum. I’m using the wrong terminology. Neutral theory is about most observed mutations being selectively neutral, not necessarily phenotypically neutral …
You claimed but didn’t prove. Praising one another’s unsupported arguments is lame.
Am I? Where? Be very specific. What happens if the dimensions are independent?
Am I? Same drill…
Really? How so? Not even asymptotic?
Huh? And once again, you offer claims without proofs. What are you afraid of?
Cartoons of marbles in jars are not facts.
Makes no sense.
Rather, polymorphisms say “natural selection” is a fantasy.
Don’t be so sure. Alleles are not exactly fair coins. Joe’s output invariably goes to 0 or 1 because the model was designed unstable. In real life polymorphism dominates.
So you think “over time… the total count difference between heads and tails will increase”? Why? Fair coin, remember?
Not always. I wrote:
“But killing the majority of one allele while sparing most of another is either a non random scenario or not possible except in the tiniest of populations and thus inconsequential.”
Here:
No it doesn’t. And if your ‘dimensions’ are NOT ‘independent’, then it would probably be worthwhile transforming them such that they are ‘independent’ and orthogonal. This assists clarity of thinking, if nothing else.
Here:
Your OPs are so densely packed with errors that I find I must restrict myself to critiquing a single paragraph.
See! That wasn’t so hard, was it? We are making progress.
Now, the wikipedia page that I linked to may be a little bit too technical for you. Would you be interested in having some help working your way through it?
Errr, it’s right there in the wikipedia page: for your one dimensional random walk, the expected distance after n steps approaches sqrt(2n/pi) as n gets large. Here’s experimental verification, where each line is actually the average of nine separate walks [because of a Monte I set up on November 3… better PRNG by far]
nonlin has some very strange ideas about stats.
Graphic:
I’ve usually seen this phrased as “confers no reproductive advantage or disadvantage.” But I can see that a mutation that is not phenotypically neutral (ie, changes the phenotype) might also be reproductively neutral.
The model is not “designed unstable”. This is the correct behaviour. In a finite population, it is inevitable that alleles eventually get lost.
In real life, mutation and migration introduce novel genetic variation. If you include this in your model, polymorphism also persists there.
See Jock’s reply.
Oh good. So populations do evolve.
Now, when they do return, how does that differ from “regression to the mean”?
Neither is your “coin” cartoon.
Exactly my point. Thanks for agreeing that your “coin model refutation” makes no sense. Never in my wildest dreams would I have imagined you agreeing that something is fundamentally wrong with your diatribe.
Nah. They say that there’s a lot of avenues for selectively neutral variation.
Nonlin mistakes randomness for evenness. In Nonlin’s imagination, if there’s a random selection from a population with 50/50 of two alleles, the alleles in the descendant population will also be 50/50, or vary very very closely to that.
Only when it suits him. Nonlin just needs species to be static.
It’s a radical departure from normal expectations on sampling – the Nonlin Sample, however small, will precisely reflect the distributions in the wider population from which it is drawn. I’d like to do a few rounds of betting on this expectation. I’d clean him out.
Entropy,
Also, fundamentally, selection could not even work without polymorphism. No allele is expected to go from inception to fixation in an instant.
That’s not equating one with the other. While the population is moving on many dimensions simultaneously, I just focus on one dimension for simplicity.
Because…?
OK. Let’s not complicate unnecessarily.
All that’s saying is that streaks end. You’re reading what’s not there.
Huh? Exponential decreases are asymptotic so there’s no need to be specific.
Hold on a sec. Say I flipped 100 times and have 55H/45T for delta100 = 10. Now I will flip another 1000 times. What is the expected total after 1100 flips? Is it not 555H/545T for a delta1100 = 10? My expectation is that delta will not increase. QED.
It’s obvious your line wanders more the more coin flips you have (and that’s what they say), but Flint’s comment fits a different scenario. The one I describe above.
See? Now you add a weird constraint: “finite population”. Are you saying: “in finite resource environments like the Galapagos diversity disappears”? I’d like some evidence.
Are you saying polymorphism is only sustained by mutation and migration? Proof? And we have the inherited and very persistent founder effect as well as genetic diseases. Not from mutation and not from migration. And let’s look at the ABO blood system. Which one is “deleterious” and will therefore disappear and how many mutation cases do you know?
They do not. You equate allele frequency changes to “evolution”, but that’s not right. Dinosaurs can’t grow wings just because some allele frequency changes.
One can only disprove a statistics claim with statistics… and then with the real life evidence presented in the other paragraphs.
Speak for yourself.
Let’s not get into “species”. Or let’s do it some other time.
You too. Speak for yourself.
This would be the case if one of the allele were superior to the others. You know? One “evolves” and the other “devolves”. But that’s not what we see. We see stasis (or at least no “evolution” trend). Remember Mendel’s pea?
If you are saying that the expectation of the number of heads after 1100 independent tosses of a fair coin, the first 100 tosses being 55 Heads, will be 555, that is correct. The more exact figures are that 0.025225018178 of the time the number of heads will exactly equal 555, 0.487387490911 of the time the number will exceed that, and 0.487387490911 of the time it will be less than that. Which is not quite the same as saying we “expect that” delta will not increase — it does so quite often, almost 49% of the time.
And of course tossing a fair coin independently more and more times is not a sensible model of genetic drift, for reasons I explained earlier and which Alan Fox has explained in his separate post.
So you do want help being walked through the wikipedia entry on random walks. Okay, I’ll have a go.
For a walk in one dimension, with Heads = +1 and Tails = -1, then the expected value after n steps is 0.
Always.
We write this as E(Sn) = 0
Always, for all n.
However, what we are all interested in, is how far the walk is from the origin. That is the expected value of the absolute value of Sn.
We write this as E(|Sn|) = {complicated equations that I cannot be faffed rendering in Latex, read the wiki page, ffs}.
As n gets large, E(|Sn|) approaches sqrt(2n/pi), err, asymptotically.
So, in your example, after 100 tosses, the expected total = 0. As ever.
But the expected distance from the origin is just under 4.
And after 1100 tosses, the expected total is 0. (In your example, if you know that the total was 10 after 100 tosses, then, as you correctly note, the expected total will be 10, for all future n). BUT, after 1100 tosses the expected distance from the origin is over 13. In your example, its over 15!
Here’s a way of thinking about it graphically:
Imagine that bell-shaped curve. It is centered on the origin, (score=0).
As n gets larger, the distribution spreads out more (this is actually the part that you fail to understand).
For coin-tossing, the distribution is symmetrical. The expected value for the total score is ALWAYS zero.
Now, instead of thinking about the total score, we want to think about the distance from the origin. So, make a fold down the axis of symmetry and fold the distribution over on itself. See how the expected distance from the origin is always a positive value? And (this you don’t get) as n increases, the expected distance increases. Always. With sqrt(n).
Thus flint’s intuition is correct.
And a well-known mathematical result.
QED, indeed.
You didn’t use statistics, you used a cartoon scenario about coin tosses. However, don’t forget that you already agreed that your cartoon doesn’t work for the purpose you wanted it for anyway.
I call’em as I see them. Mistaking random for evenness is the only reason why you would be unable to understand the results of those simulations and imagine that your non-model, but, rather, imaginary exercise of coin toss, is enough to “prove/disprove” whatever you thought it proved/disproved.
DNA_Jock,
Oops. I modeled a H:+0.5 T:-0.5 walk. I will let nonlin point out where that introduces errors into my analysis…
Nonlin.org,
Well, that’s kind of implicit in the ‘selection’ part. If there’s no such difference, there’s no selection. But what would ‘selection’ even mean if there were no variation to be subject to it?
So Mendel showed that wrinkliness and smoothness were static in the population? News to me. You don’t see population change by observing it for ten minutes.
That your understanding of genetics starts and stops with Mendel is a bit of a handicap, too.
LOL! You believe finite population to be a “weird constraint”? Where were you planning on storing your infinitely large populations, laddy?
Well, no. There’s balancing selection as well.
Founder effects result from migration and most genetic diseases persist because of recurrent mutation. Nice going!
First, your use of the word “deleterious” shows that you do not understand why genetic drift occurs. Secondly, the B-allele was lost from Native American populations, probably because of a founder effect. The indigenous population of South America was as good as fixed for the 0-bloodtype. The A and B alleles were introduced by European migrants. Finally, concerning mutation: Do you really believe the 0-allele which results from loss-of-function mutations, to result from a single mutation event that will never ever happen again?
Yes, it is. The most common definition of evolution you’ll find is a change in the heritable characteristics / genetic composition / allele frequency of a population. That should appeal to someone who likes to argue by dictionary.
Well, they did. Or do you believe the difference between winged and non-winged dinosaurs was not associated with any genomic differences?
Then that settles it.
That was not the question. If the question had been about the probability distribution… then yes.
Right or wrong, your model for “genetic drift” if not important if restricted to allele frequencies. But when it is presented as a “Neutral Evolution” scenario, that’s when we have a problem.
Maybe that’s what you are interested in. I am interested in whether “evolution” can be an outcome of randomness or not. And it can’t.
You do? I’m interested in “evolution”. Is it happening by drift? That’s why I conclude:
“And if a significant bias persists over the long term, then the coin cannot possibly be ‘fair’ “.
Obviously, the more samples you draw, the more the nominal delta(H-T) can be. So when I say:
“And over a much longer time, the realized events still sum up to an average of “no change” ”
This – as always – needs to be normalized (which you keep forgetting). Oh, look: as n goes up, sqrt(n)/n goes down. How cool is that?
So in the end what are we looking at? Some allele frequency change? How the heck will that do “evolution”? Hence “biologic drift” in the title.
We see the difference, but we don’t see any selection. That’s the beauty.
Ten minutes? That’s how many pea generations?
Joe modeled a population of 1000. That is indeed a “weird constraint”. Dying populations are bigger than that.
Haha. There’s no end to the fantasy, is it? Did you read par. 7?
Read again.
Cool story, bro. But which one is “deleterious” again? Are they all equally “fit” and now we’re just waiting for drift to do the deed? A miracle of equality?
If that were all “evolution” was about, then we wouldn’t have much to argue about. But what do you do with the old sexists, racists, and eugenists? Darwin&Co? They didn’t do “genetic composition”.
You mean dragons? “Allele frequency”, eh? Not… “Fillet of a fenny snake, In the cauldron boil and bake; Eye of newt and toe of frog, Wool of bat and tongue of dog, Adder’s fork and blind-worm’s sting, Lizard’s leg and owlet’s wing, For a charm of powerful trouble, Like a hell-broth boil and bubble.”???
This is actually true for large populations. Confirms the stopped clock hypothesis.
Nonlin doubles down. 😉
There’s a reason the Neutral Theory is known as the Neutral Theory.
Nonlin.org,
Not really any kind of rebuttal, except in the sense of being a statement in the space below mine, as is the convention.
0.00001902587. Why do you ask?
I win for the millionth time!!!!
Naah. ‘Fraid not. This is the bit that you still don’t understand. I mentioned this twice already.
Simply put : It. Does. Not.
If you really think that it “needs to be normalized”, you will have to explain what you mean by this and why this adjustment is required. You’ll have to be more precise in your language though.
But you have gotten to the bit where you noticed that as N increases, the expected distance from the origin keeps increasing.
That’s a step in the right direction [heh]. Although, as I re-read your mangled prose, you do seem to be confusing a walk with a sampling process.
Yeah, that’s right! That’s the second half of “Flint’s intuition” :
This confused you
and
You now understand that this is true, I hope…
I think nonlin.org is asking whether genetic drift (and maybe random mutation) but without natural selection, can explain all of what we see in evolution. Answer, simply, no. As anyone who has ever had a course in evolutionary biology, or read a good textbook of it, will know.
Beyond that nonlin.org is modeling something — it is not explained what — by tossing the same fair coin repeatedly, independently, and than triumphantly noting that the resulting fraction of Heads converges to 1/2.
I believe that was already known, a couple of centuries ago, And anyway nonlin.org has failed utterly to explain what its relevance to evolution is. Maybe someday they will explain it. Who knows? Who cares?
There will be genetic drift for any finite value of N.
This is getting embarassing, Nonlin. Genetic drift will occur with or without fitness differences. If there are no fitness differences, the allele with the highest frequency has the highest chance of being fixed.
Birds, Nonlin. Birds. Please don’t deny that birds have wings.
That is why you look so silly arguing regardless.
Not to worry. Since wednesday you have a new president.