
We keep hearing that Darwin is obsolete and “evolution” has turned into a real science lately. Apparently not. Here is why this relatively new theory – that was supposed to rescue “evolution” – is in fact demonstrably incorrect.
- Drift in biology is “evolution by randomness”. The ‘drift’ idea was championed by Motoo Kimura as “The Neutral Theory of Molecular Evolution”. Kimura was motivated by JBS Haldane’s dilemma: too much polymorphism exists for “natural selection” to be able to pick and chose “beneficial” genes due to gene colocation in the genome and to the propagation time in the population. Of course, Haldane’s dilemma is only due to the conflict between his nonnegotiable assumptions that “natural selection”, “fitness” and “beneficial mutations” are all real and the observable realities of biology. The ‘drift’ idea claims that random events cause allele frequencies to change over time and in extreme cases, ‘drift’ is seen eliminating or fixing certain alleles in the whole population. Drift is supposed to operate independent of “natural selection” to the extent the allele is not strongly “beneficial” or “detrimental” for its frequency to be primarily a product of “natural selection”.
- Outside of “evolution”, drift adds to nothing or is aimlessly carried by known forces. In the first case it has no effect on anything either in the short or the long term. In the second case it is explained by the carrying force and therefore is itself not an explanation of anything. Examples of driven drifts are driftwood, wind-driven snow, rain, cloud, dust, or smoke as well as aircraft moved by air currents (all these driven). Inconsequential drift examples include aimless course or random movement. In biology too, genetic drift should either be inconsequential or, if lasting changes are observed, not drift but the real driving forces behind this drift should be held responsible.
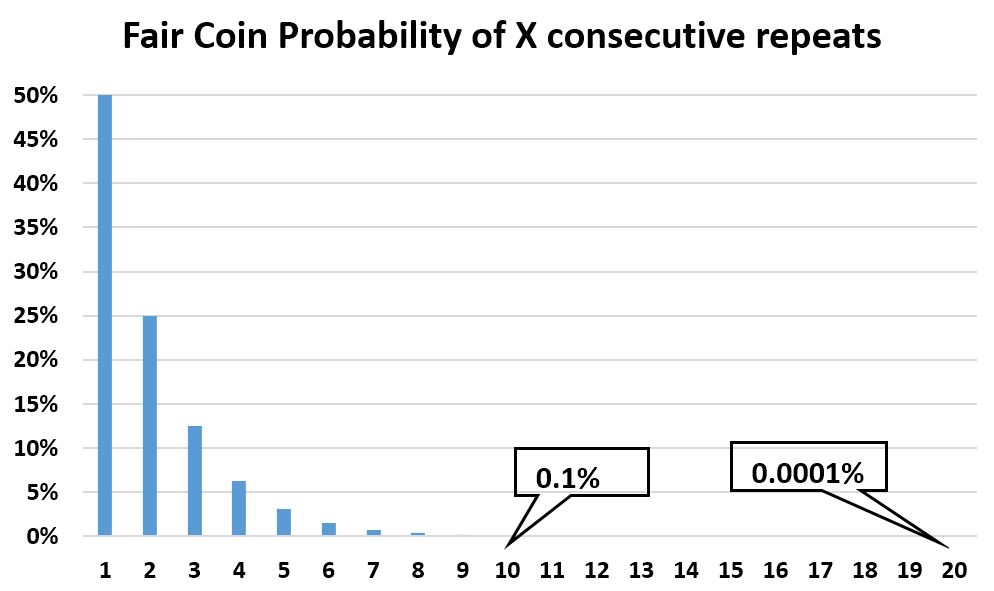
- A fair coin is a good model for inconsequential biologic drift. But how can that be, given mutations can take the population in a myriad directions, not just two (head and tails)? The fair coin model works because from a starting population, there are only two mutually exclusive directions possible on any of the myriad dimensions: towards or away from the initial value. For instance, mutations can make a second generation taller, greener, heavier, etc. But the third generation can only be even more tall, green, heavy, etc. than the original population, or less so than the second generation and therefore more like the original population. Just as with a fair coin, the output is binary: further away (call it ‘tails’) or closer to (call it ‘heads’). To prove random drift irrelevant in biology, let us consider the statistical case for drift by using a fair coin sequence. Yes, there is a statistical probability however small to see a series however large of tails (or heads), but eventually that series must end. As the chart shows, the probability of a sustained random drift decreases exponentially to nil. And over a much longer time, the realized events still sum up to an average of “no change”. This is easily verifiable experimentally. And if a significant bias persists over the long term, then the coin cannot possibly be ‘fair’.
- Population bottleneck cannot lead to “evolution”. Populations sometimes decrease dramatically due to competition, disease, predation or natural catastrophes. But killing the majority of one allele while sparing most of another is either a non random scenario or not possible except in the tiniest of populations and thus inconsequential. Post bottleneck, mutations occur and are claimed to further cause the strain to diverge from the original. But if no driving force is involved, there is no reason to believe mutations would work in a diverging direction only as the statistics discussion proves. Therefore, only a driving force, not drift can be responsible for long term changes following a bottleneck event. Alleles could indeed disappear from a population due to a series of random events, but either the population is small hence the loss is irrelevant; or, if that small population grows, a mutation will likely reintroduce the allele; or, in an already large population, the allele loss is highly unlikely due to random events not scaling up with the population size. Either way, random allele loss cannot possibly drive “evolution”.
- Founder effect also cannot lead to “evolution”. Many islands contain unique species due to having been populated by only a few organisms. Similar to bottlenecks, founder effect reduces the amount of genetic diversity in a population, meaning certain biological characteristics are overrepresented and these individuals are hypothesized to form new populations with different gene pools from the original population. But the same problem arises: the sum of mutations cannot be divergent unless a force other than drift itself causes that. If the environment weren’t drastically different, rather than diverge one would expect the new population to converge to the old strain instead given that at least that strain is known to be viable. Furthermore, if one small group found its way to the island, what prevents other small groups from repeating the trip and cancelling any such founder effect? The best explanation for the founder effect is thus that it is an ephemeral artifact and that all lasting differences are just adaptations unrelated to any random drift.
- There is more to the “dilemma” than Haldane’s take, and it’s all bad for “evolution”. A few things we know for sure: a) bottleneck populations are much more homogeneous as observed in the northern elephant seal, European bison, domestic animals, etc.; b) unique founder features (including injurious ones) persist for a long time in the founded population c) established populations (no bottleneck) are likely to be polymorphic with morphs in long term equilibrium. The theory of “evolution” predicts “beneficial” mutations taking over the population resulting in an “evolved”, better population over time. This is inconsistent with polymorphism and with the propagation of the founder’s features (some seeming “deleterious”) over time. Not only that, but normal population variability argues against “natural selection”. Because “fitness” functions cannot be calculated even in principle, let alone forecasted for any of the morphs, a hypothesized “natural selection” cannot possibly operate in biology. Furthermore, newfound populations, while adapted to their new environment, are in no way superior to their mainland cousins, even after the longest time from the split. This clearly shows “beneficial mutations” impossible. Thus, Haldane’s dilemma can easily be solved by removing his prejudiced, hopeless and disproved assumptions of “natural selection”, “fitness” and “beneficial mutations”.
- Balancing selection, an alternative to drift, fails to solve Haldane’s dilemma. The big idea of “balancing selection” is that the “fitness” of an organism with two alleles (heterozygote) is higher than the “fitness” of either allele (homozygote). Perhaps “fitness” changes in time from young to adult. Or “fitness” of a phenotype is dependent on its frequency relative to other phenotypes – rare morphs of prey are actually “fitter” due to predators concentrating on the more frequent morphs. However, there is no way to forecast anything of this sort or even to explain with any confidence the frequencies of these alleles. An elusive, fluid “fitness” attempting to explain all ends up explaining nothing, therefore it is a redundant hypothesis. And why would “fitness” changes in time? Why wouldn’t a morph “evolve” to do away with this temporal weakness? And what should the frequency and number of alleles be? Again, no rhyme or reason. Take ABO blood system. Why not more or fewer types? Why these frequencies? And why the current distribution? These are not just unanswered; they are unanswerable question with any degree of confidence.
- In conclusion, both drift and “natural selection” are disproven by polymorphism. As seen, drift in itself is not an explanation. All bottleneck drift “examples” are of theoretical cartoonish type (colored balls/ rabbits/ bugs). Some founder effect examples are real but inconsequential (northern elephant seal and Amish polydactyly) as clearly none of these leads to any “evolution”. Meanwhile, “natural selection” is perplexed and for sure inexistent as “fitness” is nowhere observable let alone quantifiable, therefore also a chimera.
Links:
https://en.wikipedia.org/wiki/Haldane%27s_dilemma
https://en.wikipedia.org/wiki/Neutral_theory_of_molecular_evolution
https://en.wikipedia.org/wiki/Genetic_divergence
https://en.wikipedia.org/wiki/Balancing_selection
https://en.wikipedia.org/wiki/Heterozygote_advantage
https://en.wikipedia.org/wiki/Polymorphism_(biology)
https://en.wikipedia.org/wiki/List_of_polymorphisms#Ants
https://en.wikipedia.org/wiki/Heterozygote_advantage#Cystic_fibrosis
https://sciencing.com/difference-between-natural-selection-descent-modification-17942.html
Is this the first time you try accessing an independent site? Because every time I’m in the EUSSR and surf the net, I wonder who are the imbeciles that elected the imbeciles that came up with something so odious. You should complain to the Galactic Senate and Senator Palpatine in particular. I am sure he will do something to you. Coming up in the EUSSR, mandatory euthanasia age “to save the planet”. But then again, the Galactic Senate complete with Palpatine clones is taking over the US too. What’s the name of that remote little planet again?
I see you’ll continue to deny in the face of the clearest evidence. And you “expected something”, but got derailed by “a bit of noise”. Expect more of that from now on. Haha.
At least you tried. But take a look again at Allan’s comment:
“If you put bacteria in their trillions into a vessel, then continuously remove a small fraction while topping up the medium to retain constant volume, you end up with a pure strain descended from just one original cell.”
I see a number of MAJOR and MEANINGFUL differences. Now, will you try again taking those into account?
What could “expected explanation” possibly mean in this context? What about “best fit reality”?
You are attempting to dispute this statement “The expected distance from the starting point always increases as time passes.” So far, you have offered zero evidence, merely displayed your ignorance.
Well, in the bacteria example, more than one member is removed at a time. So that’s going to speed things up. But it is a big bag, containing trillions of members, so that’s going to slow things down. It is still the case that, in the future, all of the members in the vessel will eventually be the descendants of a single current member.
Seven years ago, Allan covered the math involved and many of us ran sims to demonstrate this result. Check out Lizzie’s cool gif.
Still not responsible for your Tortucan ignorance.
Run a sim to prove us wrong. But we will insist on you providing your code.
I am disputing. Not “attempting”. Yours is a clearly false statement which you now deny. Even though you already agreed on the 100/1000 example I provided.
Yes, those are meaningful differences. Is that all you see? I mean it’s all in front of you. Major and meaningful. No, I am not talking about Allan eating the bacteria removed.
Btw, I asked:
If you do this experiment in identical vessels under identical conditions, could it be that in one vessel you end up with A and the other with B? And how do you know for sure this is drift and not something else? It could be some form of “majority rule”.
Let me add to that: “how long (generations) does it take on average to end up with a pure strain from a 50/50 distribution?”
Oh nonlin, the statement “The expected distance from the starting point always increases as time passes.” is entirely correct. If you think it is false, prove it!
The fact that you think this is inconsistent with the statement that the expected score is always zero, just means that you do not understand the difference between E(Sn) and E(|Sn|). Hey, I tried.
DNA_Jock,
I already did several times. You agreed and then reneged. End of story.
Tomorrow I will have fun explaining you why the m&m story is extra cretinoid. Unless you can figure out yourself. Can you?
Can’t be much trouble to copy and paste then. You know how to do that?
For someone using a pseudonym online, you care remarkably little about your personal data.
Nonlin.org,
There are differences, for sure. In the bacterial case, additional copies are provided by replication; in the ‘m&m’ example – because candy does not replicate – an analogue needed to be provided to do the same thing. The bacterial case is evidently closer to the situation pertaining in a biological population ‘in the wild’.
Nonetheless, any situation in which there is non-discriminatory removal and replication of individuals in a population will give the counterintuitive result that the population will tend to become less and less varied in time – alleles, or their analogues, are lost, meaning that one of the originals will come to dominate.
EUSSR! It’s cute when people think they’re making a devastating political point by letter arrangement – especially when they’re parroting it. It’s also the 4th Reich, apparently. 🤔
Works well with teachers’ names, too. Try it!
There’s no need to explain. Merely provide your own simulation and source code.
OMagain,
Nonlin thinks that each generation should consist of random samples from a 50/50 thing, regardless of what each generation got. Nonlin doesn’t understand the difference(s) between her/his phenotypic “model” and what genetic models are about. So, any simulation Nonlin might try, will consist of sampling from 50/50 populations each time. Explanations cannot reach Nonlin’s brain.
(Well, I suspect explanations reach Nonlin’s brain sooner or later, but Nonlin prefers to pretend that they didn’t, because Nonlin thinks you don’t know the difference(s) either. To Nonlin it’s about “winning”, not about being right.)
The fallacy being of course that you equate pestering with protection.
Allan Miller,
I notice you don’t answer any of my questions and don’t bother to link to any experimental evidence. Therefore I wonder if your bacterial example is real.
A fucked up place for sure.
Now, what’s wrong with the M&M story? The M&Ms are fine, but interpreting that as “evolution” is the cretinoid part.
The system has only one metastable point (equal numbers of each color) and [however many colors] stable equilibrium points. Once one of those stable points is reached, there’s no escape. End of story. It doesn’t matter how many times it returns to the metastable equilibrium point or how long it wanders, it will eventually reach one of the stable equilibrium points because the probability to get there is more than zero. No mystery, no magic, and most definitely no “evolution” connection.
Well, the M&M example was intended to illustrate that the [however many colors] stable conditions (that is, all the same color, whatever color it might be) was inevitably reached, sooner or later, without any actual selection taking place. As I understood it, you start with a bucket containing lots of different colors. You grab one at random and eat it without looking at it. Then you grab another at random, match it with one from an outside bag (same color), and put those two back. This keeps the total number constant, but removes a random M&M and duplicates some other random M&M.
The important thing about this process is, it shows how some characteristic or feature can reach fixation in a population (that is, every member of the population has it) even though that feature (which particular color) couldn’t be predicted at the start, and ended up possessed by every M&M even though no color selection had been done.
You are entirely correct in saying once they’re all the same color, there’s no escape. You’re also correct in saying the probability of reaching the all-same-color state is non-zero and therefore sure to happen. What you miss is, this is an illustration of how genetic drift actually works, and genetic drift is one of the several known mechanisms by which evolution happens.
Oh the irony.
What Flint said.
I wonder, why did you call equal proportions of each color a “metastable” point? It is no such thing. The only stable points are the absorbing states where one colour is fixed.
Corneel,
Much as I hesitate to try to explain what nonlin might mean (heh), I suspect that in his parlance equal proportions of each color is ‘metastable’. Just by symmetry. He simply failed to notice that any proportion of any number of colors greater than one is also ‘metastable’, in his parlance.
But I am sure he will clarify.
Right!
Only it doesn’t work in large enough populations (the only ones that matter). In real life that is. As polymorphism proves. So no, life is not M&Ms.
Even if it did, how does allele frequency link to “evolution”? I asked before: is the human just a snail with different alleles?
Let’s say ‘top diversity’. I haven’t run the numbers but it might also be the highest probability of “no change”. I knew someone will hang on to that little straw for dear life 🙂
Talk about “clarify”. I notice neither Allan nor anyone else is willing to answer my questions re “bacteria instead of M&Ms”. Should I assume that was all fake science?
Yes, it works in any population of finite size. You are in denial again.
Again you assume that what you want to be true is actually the case. No, the point with equal proportions has the highest variance and therefore it has the lowest probability of “no change”.
What “it” are you talking about?
1. The M&M experiment predicts “no polymorphism” (or at minimum an elimination trend). Is that true? In life, large populations, not in M&Ms! NO! It isn’t.
2. And EVEN IF fixation were true (in large populations, not a handful of M&Ms), that would work AGAINST drift, wouldn’t it? We have confirmed that once you reach those stable points, there’s no going back in the M&M experiment.
3. Furthermore, EVEN IF alleles drift could be confirmed “random” as opposed to driven by other factors (it can’t be proven as explained!), what is the “evolution” angle? What has allele frequency to do with “evolution”? Nothing! That’s what.
Like I said, I haven’t run the numbers. One way or another, does that prove “evolution” in any way, shape, or form? NO! It doesn’t.
Folks, give up, you can’t win this argument. Because any simple and clear demonstration you give of evolutionary processes will be rejected by nonlin.org as “not evolution”.
You made the mistake of arguing with The Decider, who is the final authority on that.
“The M&M experiment” is an educational tool to demonstrate how genetic drift in isolation affects the genetic composition of a population. Its value as a predictive tool is limited by the fact that, among other things, no new “alleles” are being introduced by mutation or migration. If you want that you can simulate that. Here is how you do it:
1) With every step of the process, cast two dice.
2) When you throw snake eyes, add in a mutation step:
3) Grab one additional M&M at random from the bag and eat it without looking at it. Replace it with an M&M from another bag that has equal numbers of all four colours. This simulates the influx of new alleles by mutation or migration from a large outside population.
4) Continue
Even though the colour frequencies will still be continuously changing, you will never again see a population trapped in an absorbing state. If you want this simulation to be even more realistic, occasionally introduce new colours into the mix, to simulate the mutation to novel alleles.
Now please stop using that silly “no polymorphism” argument.
Did you study the Buri experiment? Did you notice that the three genotypes correspond to distinct eye colour phenotypes?
It certainly doesn’t. What it does show is how your mind works. Think about it. Why did you feel compelled to call the point with equal proportions a “metastable point”? Why, just because you desperately want it to be. I bet that you never checked whether it was actually true, or even bothered to look up what a “metastable” point is.
It’s easy to win this argument. The hard part is convincing Nonlin that he has, in fact, lost.
But you are right of course. Time to bail out again.
What Corneel said. The various computer programs to simulate drift are models. In the absence of new mutations and in the absence of purifying selection, where there are competing alleles for a particular locus, one allele will eventually fix with the loss of the others. That’s what the models show.
I dunno, he must have some intuition that his ideas have no value. He’s touted them at various other sites and received exactly the same feedback as he gets here. Not even some of our other evolution skeptics have said anything remotely resembling “Your ideas are intriguing to me, and I wish to subscribe to your newsletter.”
I can claim the same about you, can’t I? You don’t seem to show any thinking flexibility.
That’s fine. But that’s not the issue, is it? I even numbered the issues to make it easy for you to address 🙂
And once you mess up with your model, you might as well give it up. Why would anyone think the M&M exercise relevant to biology in the first place?
In real life we see polymorphisms – some looks stable, some may be changing over time. But there is no way to trace whatever happens to drift as opposed to other factors including mutations and migration.
No. You’ll have to describe and make your case for whatever conclusion you want right here. Then, if I see any issues, I might go read to doublecheck your analysis.
Give it up on the psychoanalysis fraud. Well, I guess “evolution” goes hand in hand with dr. Fraud, astrology, palm reading, phrenology and other pseudo-sciences. If I were so inclined, I might say: “this does show how your mind works” 🙂
Once again, the biggest question is: why is allele frequency “evolution”?!? Consider a population with black and white organisms. Which one would you say is more “evolved”, the black or the white? What would Darwin say about this? Galton? Haha.
Nope. You cannot.
You, on the other hand …. hum! None whatsoever! Even when you contradict yourself, you’d rather remain silent about it than show some “thinking flexibility.”
🤣
Because it’s an illustration showing that selection is not necessary for getting to a single color, contrary to “intuition.” Just like alleles can be fixed in a population without it being necessarily due to the allele being advantageous.
It’s a tad hypocritical to complain about an M&M illustration, when you have that poorly thought “coin model” in the OP. Beam in thy own eye and all that.
There’s many tools available for checking which is which. They require a lot of patience and data. Joe has online-available books that explain some of these tools, but understanding them requires some reading for comprehension capabilities that you lack. Your ignorance is a reflexion of your ignorance, not of lack of approaches and analysis tools.
Because evolution is about heritable characteristics, which are represented by allele frequencies in wide terms. Because, as populations diverge with time, their allele frequencies diverge.
Neither. The concept obviously escapes your poorly prepared mind. Polymorphisms inside populations correspond with allele frequencies within the population. Each generation, the whole population would be “more evolved” than the prior one, given that its allele frequencies change each generation, with new alleles appearing, some old ones disappearing, etc.
That evolution has become a very interesting and fruitful field of research. That it’s amazing how much about evolution can be captured in such a simple concept as changes in allele frequencies in populations.
And you didn’t even notice that all “numbers” were addressed.
Look, it is obvious that you have no interest in learning about my position and since you refuse to tell me about yours, I frankly see no reason to continue. I will be following Joe’s advice and will be leaving this discussion. The floor is yours.
Perhaps you need to define “evolution” and explain why you have it scare-quoted….
Were they? Let’s see:
1. “With every step of the process, cast two dice.” is supposed to account for polymorphism. The problems are a) it’s a different model so why not start with this instead? b) it still converges only slower to the same stable points (or not at all). c) why is this a good model, and what is it proving?
2. When you throw snake eyes, add in a mutation step vs “doesn’t fixation works AGAINST drift?”. Add enough mutations and model doesn’t converge anymore. So model loses all meaning. And this still does not answer the question.
3. This simulates the influx of new alleles by mutation or migration from a large outside population. is supposed to address “What has allele frequency to do with “evolution”?” It doesn’t.
Or perhaps this is the answer to #3: “Did you notice that the three genotypes correspond to distinct eye colour phenotypes?” But all this does is point to [sometimes] link from genotype to phenotype. OK? And the “evolution” angle is…? Think of little monkey, what alleles would you change in little monkey to make a human out of it?!? Impossible! That’s not what alleles do.
Little monkey has an opinion of his own (from the operating table no less): something foggy about “represented” and “wide terms”. Maybe you can translate monkey talk? Of course it’s all from “evolution” to alleles. But I asked from alleles to “evolution”.
To conclude, between irrelevance to real life and having to modify it beyond recognition to match real life a tiny bit, the M&M model is bogus. Of course, we could talk about real life bacteria instead, if Allan could backup his claims.
Joe likes to play in his own sand box where no one challenges him because he can suppress dissent. Occasionally, he exposes himself here, where he ends up on the losing end, only to go back to his echo box where he distorts the discussion to make himself a winner.
But why do you expose yourself in the first place? Lots of people are afraid – rightfully so – that allowing any critical analysis of “evolution” may destroy the sand castle. Hence the vicious propaganda war against all criticism. What’s up with your bipolar approach?
The answer to “3” is that your model is absurd. But you’re too much of an illiterate to get it.
I love the “[sometimes],” for fear that you’d be taken to admit a relationship between genetics and the organisms charactaristics. No sir! There’s no genetics, not in Nonlin’s world, where Nonlin looks like monkey mama and monkey papa because god build Nonlin magically in mama’s womb and made Nonlin similar to the monkey parents. Not because of some bullshit about inherited genes.
That alleles can get fixed in a population against naïve “probability” intuitions, such as those of a little monkey called Nonlin. Isn’t this thread about genetic drift despite the wrong word in the title? Have you forgotten what this was about Nonlin? Is this one more instance of your inability to agree with yourself?
Of course not. Alleles cannot think about what alleles to change to turn you into a human. The question is ill-posed, no surprise you being a little illiterate monkey. The proper question is which alleles are behind the divergence from an ancestral monkey to you on one side, and to humans on the other.
The answer? I’d ask you for funds to figure it out, but you think that answering your questions is equivalent to leeching out the tax payers, so I’ll have to say the answers must be somewhere in the millions upon millions of alleles that differ between you, little monkey, and any human. Many of them have diverged by genetic drift. Some under selection. Some researchers are investigating which ones show selection-related marks, between chimps and humans, but not between little monkeys like yourself and humans, since that distance is higher, thus involving many more drift-fixed alleles than humans vs chimps. I guess we’ll have to wait for the chimp research to get some answer.
You cannot help behaving like the little idiot that you are. What makes you think that you represent a challenge to anybody when you cannot deal with simple concepts? I have seen Joe being authentically challenged by people who do understand what he’s talking about. For example in scientific conferences. Those are discussions where we can learn something. Discussions with you are about whether you’ll be able to understand the simplest foundations, whether we’ll be able to help you out of the most obvious of your mistaken notions. But scientific challenges? Perhaps in your dreams.
The only exposure Joe suffers here is to your childish insults and mentality. You’re light-years away from being able to challenge Joe, or me, or just anybody here, on anything but getting you to understand simple words. You’re a simpleton Nonlin. An absolute idiot. A poster child for the Dunning/Kruger effect. Nothing else.
Sorry. But someone had to give you the sad news. You’re just a petulant buffoon.
In a low IQ display, little angry monkey has no business other than squatting on other peoples’ threads and flinging poo. Will the owner of this nasty monkey come pick it up before animal control is called.
Someone named Joe must be the owner. Little monkey keeps calling your name, Joe. If you read this, come pick up your nasty little monkey.
Thanks,
If you don’t want people responding to your writings, post them elsewhere where you can control who posts what.
Says the poor idiot who couldn’t get the message, and responded with the very same immaturity and idiocy that got the poor idiot in trouble in the first place.
Says the poor idiot who flings poo towards professionals whose mistake has been to try and get the very same poor idiot to understand basic concepts.
Careful, they might take you away, since you’re the only little monkey around here.
As I said, nothing but childish insults and childish mentality from you. Thanks for your commitment to confirm what I said, but there was no need.
Entropy,
He’s the black Knight. Very soon, he’ll bite our legs off.
Alan Fox,
The spoiled brat thinks that everybody should respond in admiration to her every word, just because her parents smell her shit and smile.
Oh no! Now it’s a troop. Oh well.
Nonlin.org,
You cannot get a clue, can you? Maybe next time I should keep all subtleties out.
🤣