
As shown repeatedly, “Nothing in Evolution Makes Sense. Period.” Not natural selection, gradualism, human evolution, UCD, tree of life, etc. And just to confirm, let’s look at another one of the nonsensical concepts of “evolution”.
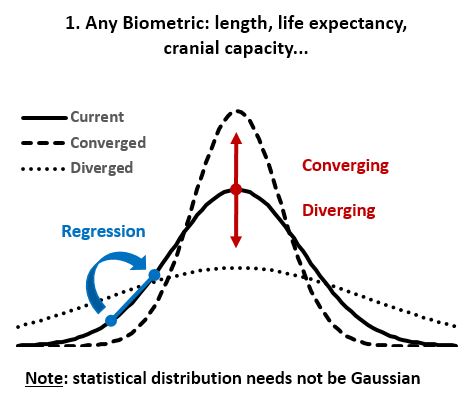

- ‘Divergence of character’ (character displacement or sympatric speciation) postulates: “during the incessant struggle of all species to increase in numbers, the more diversified these descendants become, the better will be their chance of succeeding in the battle of life. Thus the small differences distinguishing varieties of the same species, will steadily tend to increase till they come to equal the greater differences between species of the same genus, or even of distinct genera” (Darwin 1859). Sympatric speciation is hypothesized as “the evolution of a new species from a surviving ancestral species while both continue to inhabit the same geographic region”.
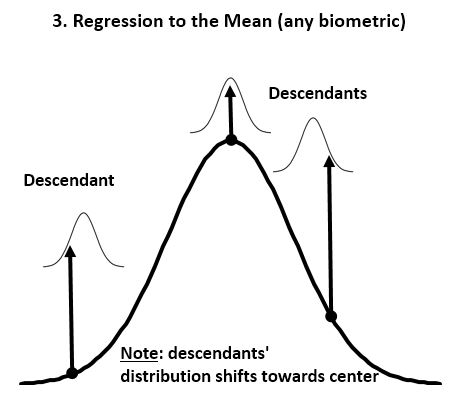
- ‘Regression to the mean’ is the biological law that overrules passive ‘Divergence of Character’. Any homogeneous population can be sorted statistically on various biologic metrics, usually resulting in a Gaussian (normal) distribution that is conserved over time in the absence of major environmental changes (as Mendel first showed; Fig 1&2). ‘Regression to the mean’ is thus the rule that causes the progeny of extreme individuals to be less extreme than their parents. Two outstanding tall parents will have statistically shorter children, and the progeny of the most and least intelligent/strong/aggressive/attractive/etc. will be more average than the parent. Many of the extremes have no descendants at all due to their limitations, and thus their “contribution” to the next generation is simply the average individual.
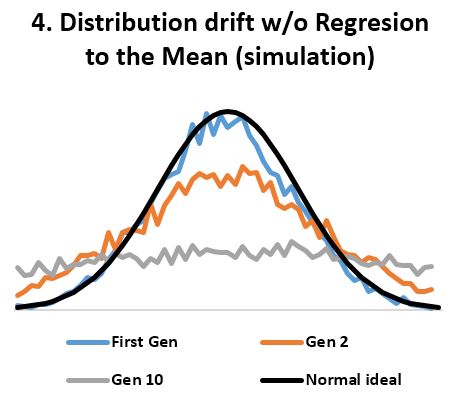
- In stable environments, population variability is extremely well conserved from generation to generation (Fig 3) as documented by the fossil and many other records. ‘Regression to the mean’ is thus a mathematical necessity without which a passive ‘divergence of character’ would be observed in very few generations (Fig 4). ‘Regression to the mean’ mechanism is incredibly accurate and allows for conservation of traits over thousands upon thousands of generations as observed. Scientists were rightfully surprised that ancient bacteria and many other fossils as well as mummified organisms including cats and monkeys are indistinguishable from their contemporary descendants. At a minimum, the number of organisms that show remarkable stability over long periods (living fossils) invalidate the ‘General Divergence’ theory. Does a limited, ‘Special Divergence’ hypothesis still make sense?
- Observed long term regression is highly unexpected and contrary to ‘divergence of character’ and ‘drift’ hypotheses. ‘Regression to the mean’ operates in the longest term observed, whenever environmental conditions are restored following significant changes that led to adaptive mutations. Most – if not all – organisms are endowed with a limited ‘plasticity’ trait that allows them to retain adaptive characteristics for generations. And yet, when the stimulus that caused the adaptation disappears, these organisms regress rather than maintaining those adaptive traits or accumulating even more diverging ones. Darwin’s finches, the peppered moth, antibiotic resistant bacteria and the domesticated plants & animals – all these and more have been observed to regress to the old mean when the adaptive stressor is removed, thus disproving even the limited, ‘Special Divergence’ hypothesis. These are not coincidences! The regression can happen over a few generations as in most epigenetic changes, many generations, and even the indefinite future if the adaptive stimulus is maintained (such as in domestication). Biologic variability can be compared to a loaded spring – the more it stretches, the harder the pull back (regression to the mean) and the more fragile is the extreme variant population. Domesticated plants and animals show that crossbreeds are resilient, while pure breeds are fragile showing that extinction of the extremes is the default outcome that promotes the ‘regression to the mean’ of the extended population.
- Adaptation neither demands not implies divergence in any way. What about the ‘adaptive radiation’ seen in Darwin’s finches, the cichlids of the African Great Lakes, and others? Is this not ‘divergence of character’? No. The driving force in all these and more is adaptation, not divergence even if “evolution” were true. Organisms just seek survival and, if their built-in yet limited plasticity matches the environmental challenges, these populations survive as variants. Otherwise, they simply go extinct like many others before. The new traits are not ‘divergent’ as shown by all known cases of reversals (as discussed) and none of further divergence when the adaptive stressor is removed. If ‘divergence of character’ were true, adaptive plasticity traits would be cumulative and sticky even after the adaptive stressor was removed, and the more extreme variants would be at least as resilient as the mean. Furthermore, experiments would show increasing variability over time in all research organisms and even more so in the short lived ones like bacteria. There would not be any distinct “species” and organisms would freely undergo metamorphosis (transmutation) into one another. Differential survival and randomness would eliminate all but the “best adapted” allele, therefore the Mendelian conservation of alleles would not be observed. Yet none of these are happening, thus falsifying the ‘divergence of character’ hypothesis.
- Adaptation is “fast and done”, “do or die” by necessity, unlike the supposed “slow and ongoing” ‘divergence of character’. If adaptation is not fast enough, the population simply goes extinct as many others did. The cichlids of Lake Victoria had less than 15,000 years to adapt and are as diverse if not more so than the cichlids in the other, much older African Great Lakes. But they do not need even that much time as the newer aquarium varieties obtained in a few generations show. Most likely, cichlids variants have come and gone throughout the history of all African Great Lakes in short cycles of adaptation. And that is why the cichlid biodiversity difference between a few years (Lake Victoria) and millions of years (other African Great Lakes) is unremarkable. The only remarkable fact is that cichlids have a predominantly Gondwanan distribution showing that in 180+ mil years, they did not adapt to ocean living despite their otherwise high adaptability. This clearly shows the limitations of adaptability and makes it an unlikely substitute to ‘divergence of character’. Darwin’s finches, peppered moths, bacteria, and many other also adapt fast or die as observed. And when the stimulus disappears, they revert just as quickly, and later readapt to whatever new stimulus they face or simply die out trying as confirmed. It is a very good thing ‘divergence of character’ is false, or else antibiotic resistant bacteria and other superbugs would have killed mankind by now as “evolution” falsely predicted.
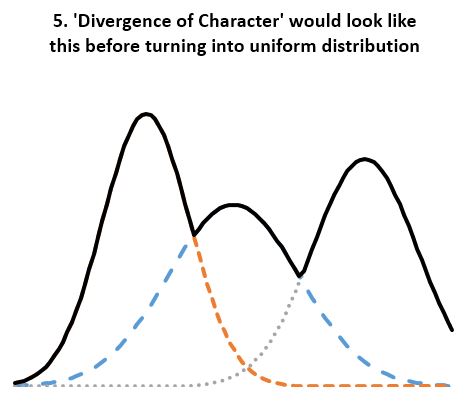
- Statistical evidence refutes ‘divergence of character’. According to the theory, “when organisms compete for scarce resources, natural selection should favor those individuals that are least like their competitors”. And since organisms always “compete for scarce resources”, the least average members of a homogenous population should always be favored by “natural selection”. If so, the well known normal distribution of any organism dimension (length, height, weight, etc.) should always be under pressure to change. We should see groups of “least like” the average form second, third, and so on normal distributions of their own, thus reshaping the original normal distribution into a composite distribution with several peaks and valleys as in Fig 5. And even that should not be adequate, as any concentration of similar individuals would be disadvantaged according to the ‘Divergence of character’ hypothesis, thus leading to uniform distributions as in Fig 4. However, neither Fig 5 nor uniform distributions are seen in homogeneous populations. Instead, we always see normal distributions. And since we see the normal distribution maintained over arbitrary number of generations and no hint of transitioning to a uniform distribution, the ‘Divergence of character’ hypothesis must be discarded. A trend not supported by several period observations must be discarded as noise artifact. This is the case for all examples considered including Darwin’s finches, the peppered moth, antibiotic resistant bacteria, cichlids, etc. All seem somewhat supportive of the divergence hypothesis over carefully chosen periods, yet the divergence is clearly illusory over longer periods.
- Are the bear of North America not like Fig 5? Yes, but they occupy different geographic regions. They are not homogenous. Indeed, we do encounter subfamilies of organisms, that have normally distributed metrics within the subgroup yet clearly distinct from those of other subgroups. However, where these subgroups overlap, the blend is always geographic and never biologic, meaning we see fewer of one kind and more of the other when moving from one’s territory to the others’ instead of blended characteristics as ‘divergence of character’ would predict. Humans are not different “species” although various subgroups are exclusively vegan/carnivorous, white/black, extra small/large. And domesticated organisms including canids are even more diverse than humans. Are the wild cichlids, finches, mice, and others qualitatively different than humans and canids? No. Then why the different “species”, many of which, ironically, are threatened by hybridization? The unwarranted inflation of “species” that do not even meet the loosest definition of reproductive isolation has the sole purpose of perpetuating the myth of ‘divergence of character’.
- Multimodal distributions in homogenous populations are not due to ‘divergence of character’. Indeed, bimodal distributions (Fig 2) and multimodal distributions are not uncommon in homogenous populations. However, these are due to the discreteness of physics in general and biology in particular, not due to ‘divergence of character’. Male and female populations are not diverging from one another and various alleles are in long term cyclical equilibrium as shown (spring model). ‘Drift’ is often invoked as a mechanism of ‘divergence of character’. This is wrong because ‘drift’ explains nothing as it is either aimless noise or due to adaptation and environmental change. Yet, as shown, adaptation is in no way ‘divergence of character’. In addition, the stable coexistence of several distinct variants within a homogenous population shows “gradualism”, “survival of the fittest”, and “natural selection” to be false because the alleles responsible are themselves distinct (no “gradualism”), they all “survive”, and neither is “selected” for or against.
- Darwin worried about regression to the mean for the wrong reasons. Namely, if blending inheritance (Darwin laid an egg) was true, then natural selection could not be true. Darwin puzzled over this a lot, but ended up with nothing satisfactory. Then Mendel showed that inheritance is discrete, not blended. Mendelian Inheritance Tables (see Punnett squares / Hardy-Weinberg equilibrium) show “probabilistic traits conservation” and thus disproving ‘divergence of character’ (at least as byproduct of reproduction) as well as dismissing “gradualism” (another one of Darwin’s unsupported claims).
- When entire populations split, do subgroups diverge from one another? This is not how ‘divergence of character’ is supposed to work.Descendants are supposed to diversify within the homogenous population. Furthermore, populations split by environmental conditions simply adapt to the new environment and for as long as those conditions allow. Adaptation is the driving force with no ‘divergence of character’ anywhere in sight. Island biology is the most diverse because islands are isolated and have many microenvironments. However, island variants are close descendants of their original colonists, showing that no divergence ever happened. Their risk of hybridization is high, disproving the “speciation” claim. They are also fragile examples of the extreme stretched biological spring model discussed, and will likely go extinct if at all stressed and when interacting with mainland.
Summary:
1. ‘Regression to the mean’ is the biological law that overrules passive ‘Divergence of Character’
2. In stable environments, population variability is extremely well conserved from generation to generation
3. Observed long term regression is highly unexpected and contrary to ‘divergence of character’ and ‘drift’ hypotheses
4. Adaptation neither demands not implies ‘divergence of character’ in any way
5. Adaptation is “fast and done”, “do or die” by necessity, unlike the supposed “slow and ongoing” ‘divergence of character’
6. Adaptation has limited powers and is thus not a substitute for ‘divergence of character’
7. ‘Divergence of character’ hypothesis would lead to uniform rather than normal (Gaussian) distributions as observed in homogenous populations
8. A trend not supported by several period observations must be discarded as noise artifact
9. The unwarranted inflation of “species” that do not even meet the loosest definition of reproductive isolation has the sole purpose of perpetuating the myth of ‘divergence of character’
10. Multimodal distributions in homogenous populations are not due to ‘divergence of character’
11. Mendelian tables show “probabilistic traits conservation”, disproving ‘divergence of character’ (at least as byproduct of reproduction), as well as dismissing ‘gradualism’
12. Island biology proves adaptation and the biologic spring model while disproving ‘divergence of character’
13. What’s in, what’s out? IN: ‘regression to the mean’, ‘adaptation’, coexisting variants, long term stability, spring model, normal distributions. OUT: ‘divergence of character’, gradualism, drift, speciation, uniform distributions, “natural selection”, “survival of the fittest”, “evolution”.
Links:
https://ucmp.berkeley.edu/bacteria/bacteriafr.html
https://www.sciencedaily.com/releases/2019/10/191018112136.htm
https://www.ncbi.nlm.nih.gov/pmc/articles/PMC3285564/
https://www.ncbi.nlm.nih.gov/pmc/articles/PMC3352989/
http://www.galton.org/essays/1880-1889/galton-1886-jaigi-regression-stature.pdf
https://en.wikipedia.org/wiki/Character_displacement
https://en.wikipedia.org/wiki/Sympatric_speciation
https://www.bionity.com/en/encyclopedia/Character_displacement.html
https://biologydictionary.net/divergent-evolution/
https://en.wikipedia.org/wiki/Cichlid
https://biology.stackexchange.com/questions/41982/regression-to-the-mean-and-evolution
It’s probably just careless choice of language, but those statements read as inconsistent.
Anyhoo, a question you may have missed: under your proffered definition of sex – “two individuals of different types (sexes)”, how did the parents come to be different? I mean in their autosomes, I’m not looking for an account of gender. Were organisms just Created different?
Also, ISTM that the logical endpoint of the narrowing of a distribution would be no distribution at all – the eradication of all variance, at the limit. What opposes that, to retain genetically variant parents in a population? Environmental variance would still exist, but you’re surely not basing the distinction on that?
Quoting: “You can’t claim one BS based on another BS.”
Citation please. Where did you get that idea?
In my understanding, divergence of character in sympatric speciation only occurs in special circumstances, one of those being the opportunity to specialise to different niches. Otherwise, one (sub)population is likely to go extinct because of competitive exclusion.
I was responding to:
Inbreeding is extremely common in nature. If you insist on sex being the sweaty body thing between two individuals, there are still plenty of examples of species with high levels of inbreeding. Jock already mentioned C. elegans nematodes and baker’s yeast. There’s many more.
Allopatry presupposes evolution? That’s a new one.
Anyway, looking forward to your code.
I look forward to that promise being fulfilled. 😏
They do not. “Variation” is the main ingredient in “evolution”. Remember? But since “evolution” is not true…
“Not producing clones” is how sexual reproduction is different than asexual reproduction as far as outcome.
Not missed, but not a good question. You can do a mouse “Adam and Eve” experiment.
What narrowing? It seems regression and mutations are opposites. Bottom line: stasis aka “living fossils”.
Misquoting, rather.
See first paragraph. Like I said: why do I know your “evolution” better than you?
“Allopatric speciation” sure does.
This very long side discussion was at one point about clones, aka [supposedly] 100% identical DNA.
On another note: you’re not making any progress? When will you read beyond second paragraph?
Yeah, so you say. There’s no variation then. Gotcha.
You said the whole point, rather than simply ‘this is what distinguishes them’.
No, it’s an excellent question. A mouse Adam and Eve would have, at most, 4 alleles at a given locus. At most loci, fewer, otherwise their progeny would not be capable of meiosis, which depends on a high degree of homozygosity. So by mating brothers and sisters over multiple generations, you would end up with a great deal of inbreeding, and even less ‘variation’ than you started with.
If populations ‘regress to the mean’, you are talking about a distribution – a bell curve. If there is a ‘central tendency’, that bell curve will narrow – variance will get less. So the ultimate ‘stasis’ has no polymorphism, and hence no opportunity for heterozygosity in zygotes formed from ‘different’ parents. They aren’t different at all, if there’s no variation left. Yet this is not what we observe.
You made this comment in another thread:
My question is: how would you differentiate between a real S-L-O-W process and a fake one? Because I address this in paragraph 7 of this OP, but it seems you haven’t read that far.
To your claim that “evolution” is JUST “genetic changes occurring”, if that were all, we would be done right here, right now. But I smell a bait and switch. Right?
I didn’t say “there’s no variation”. Read again.
But it is “the whole point”. Why would anyone design another, more complex reproductive process to get the same exact results? It would be illogical.
Aren’t you forgetting the mutations? It happens in labs too. Without tight controls, they would get LESS inbreeding over time.
Not how it works. You misunderstand.
Don’t forget your promise!
Well, I haven’t forgotten them, but you appeared to. Mutations = evolution, which you say doesn’t happen.
Also your statement is flatly contradicted both by theory and observation. Small populations lose variation very rapidly. It is not replaced sufficiently quickly by mutation, which is why there is such concern over what amounts to inbreeding in endangered species and pedigree breeds.
So you are invoking a statistical phenomenon which doesn’t affect a future statistic? Guess I do misunderstand. 🤔
False. Mutations ≠ evolution. We do not need “evolution” to explain mutations.
We were NOT talking about small populations. Of course things will be different in THAT case. Stop moving the goal posts.
You have not contributed any meaningful thoughts to this discussion. Once you do, and I see a specific need for clarification, I will send those details to you. Meanwhile, you have the description of the model in a reply to Corneel if you want to study. Looking forward to your upcoming intelligent questions/comments.
Translation: “no, I can’t provide any evidence for my assertions.”
Nah. It’s more like “I made those graphs by hand and presented them as being from simulations.”
False. Mutations = evolution. Mutations is one of the causes of why evolution occurs. Nobody says evolution “explains mutations”. It’s the other way around. Mutations occurring for well understood chemical and structural reasons, is a cause of evolutionary change and divergence of character.
I was tempted to ask whether MS Paint was involved. Nonlin should be able to tell us what he used as raw data and his algorithm for his simulation. He should look at DiEb’s histograms on TSZ and UD comments for a clear example of how to show his work.
No, we don’t need evolution (oh, how you love your “scare quotes”!) to explain mutations, but mutations are “evolutionary”.
You don’t get much smaller than a ‘mouse Adam and Eve’. The point under discussion was how a varied population comes to be varied, and ‘mouse Adam and ‘Eve’ was your response, with an assertion that inbreeding should decrease over time, thanks to mutation. The evidence contradicts that.
You’re cavorting round the pitch like a mad thing then complaining it’s the goalposts that have moved! 😃
Corneel,
Apologies, Corneel. I haven’t been taking Nonlin seriously and, as a consequence, I did skip over some comments. I just reread your comment above and it sums up the difference between heritable and non-heritable traits and their expected distributions very clearly.
ETA: live link
Not seeing any mention of “homogeneous populations in a constant environment”. You are saying that Charles Darwin made that claim? Are you sure?
It sure does, but Jock didn’t SAY speciation. He said allopatry. You don’t presuppose evolution by modelling allopatry.
These side discussions tend to grow very long because you resist correction of even your most egregious misunderstandings. When Jock took issue with your statement that “sexual reproduction […] cannot produce clones even theoretically” he was completely right. What is it that makes you defend the smallest molehill, at the expense of demolishing every opportunity to appear even half-way reasonable?
That comment was addressed at Robert Byers, who inhabits an entirely different realm of denial than you. His position, stated in the OP, was that if we don’t observe something happening in human time-scales, then we somehow cannot establish that it could happen ever at all. KN and me had a little fun with that by listing everyday phenomena that are too slow to observe happening in real-time. but that everybody accepts.
If you cannot appreciate the ludicrosity of Robert’s argument I will have to move you above Robert Byers on my personal TSZ sillyness scale.
New favorite comment! 😀
Yup, “regression towards mediocrity” was once taken seriously as an impediment to evolutionary change, in the Victorian age. The apparent paradox was solved long before the modern synthesis arose, by Karl Pearson (of correlation coefficient fame).
I am confident that he used Excel (or similar) to generate his “target” curves (normal distributions with increasing variance) and then “hand-drew” noise onto his data values in Excel to make Figure 4 look like the results of a sim.
My reasoning:
Not Paint, because the data points line up with each other on the x axis.
Human-generated noise, because the “noise” in Figure 4 is waaaay too regular to be actual noise; it looks exactly like alternates-too-much sequences that naive humans view as being random.
I look forward to his proving me wrong with his code.
And that, folks, is how actual design is actually detected.
False. You don’t know the meaning of ‘=’.
Try ‘=>’ instead. But then it’s on par with “stars position => your mood”, but worse give ‘mood’ is a real thing unlike “evolution”.
DOES NOT. ‘Mouse Adam and Eve’ implies unrestrained growing population. Read the story of Adam and Eve! WTF?!?
He certainly didn’t say “changing environment”, so by default nothing changes. Also, in the same paragraph, “Sympatric speciation is the evolution of a new species from a surviving ancestral species while both continue to inhabit the same geographic region. ”
Reply with a phrase where ‘allopatry’ does not imply in any way “evolution”. That should be fun (whatever happened with the other fun we were going to have?). And in any case, the topic is ‘divergence of character’ as in “evolution”.
Actually, no. He conceded that sexual reproduction does not make clones: “Well they don’t have 100% the same genome. ” http://theskepticalzone.com/wp/divergence-of-character-myth/comment-page-2/#comment-266529
Add to that the fact that his examples organisms are hybrid and not classical sexual reproduction.
Why is this important? As discussed, it would indeed be great material for a new book if we found out sexual reproduction had the same outcome as asexual reproduction. Do you get it? Curious mind are always on the lookout for interesting discoveries.
You’re trying to win on technicalities. But it won’t work. When you and all others will go silent as you ALWAYS do, I will know you conceded. Which you should do out loud if you indeed seek knowledge and not just defend an untenable, irrational belief in “evolution”.
You listed “new mountains popping up” which is something we actually DO observe in human time-scale. So you defeat yourself.
And what are we supposed to take as basis of belief if not confirmed observations?
You continue to not read paragraph 7 (or anything inconvenient), so here it is:
“…And since we see the normal distribution maintained over arbitrary number of generations and no hint of transitioning to a uniform distribution, the ‘Divergence of character’ hypothesis must be discarded. A trend not supported by several period observations must be discarded as noise artifact. This is the case for all examples considered including Darwin’s finches, the peppered moth, antibiotic resistant bacteria, cichlids, etc. All seem somewhat supportive of the divergence hypothesis over carefully chosen periods, yet the divergence is clearly illusory over longer periods.”
Do you agree with the highlighted sentence which is incompatible with “evolution”? Why/not?
I see you’re dying of curiosity, so here it is. No “hand-drew”.
Obviously, first gen doesn’t have the offsetting “A3 ” you see in the formula cell.
Columns L:Q show binning.
Of course, this model proves only 1. in the summary of this OP. So when will you address all other points demonstrated???
Summary:
1. ‘Regression to the mean’ is the biological law that overrules passive ‘Divergence of Character’
2. In stable environments, population variability is extremely well conserved from generation to generation
3. Observed long term regression is highly unexpected and contrary to ‘divergence of character’ and ‘drift’ hypotheses
4. Adaptation neither demands not implies ‘divergence of character’ in any way
5. Adaptation is “fast and done”, “do or die” by necessity, unlike the supposed “slow and ongoing” ‘divergence of character’
6. Adaptation has limited powers and is thus not a substitute for ‘divergence of character’
7. ‘Divergence of character’ hypothesis would lead to uniform rather than normal (Gaussian) distributions as observed in homogenous populations
8. A trend not supported by several period observations must be discarded as noise artifact
9. The unwarranted inflation of “species” that do not even meet the loosest definition of reproductive isolation has the sole purpose of perpetuating the myth of ‘divergence of character’
10. Multimodal distributions in homogenous populations are not due to ‘divergence of character’
11. Mendelian tables show “probabilistic traits conservation”, disproving ‘divergence of character’ (at least as byproduct of reproduction), as well as dismissing ‘gradualism’
12. Island biology proves adaptation and the biologic spring model while disproving ‘divergence of character’
13. What’s in, what’s out? IN: ‘regression to the mean’, ‘adaptation’, coexisting variants, long term stability, spring model, normal distributions. OUT: ‘divergence of character’, gradualism, drift, speciation, uniform distributions, “natural selection”, “survival of the fittest”, “evolution”.
Where’s the proof? And EXPLAIN. Don’t just link to somewhere in the ether!
My apologies, nonlin. You merely picked a really bad example of your “noise” for the OP.
So you have modeled drift. Notice how the Std dev increases with the square root of the generation number. That’s a fairly well known result.
How are you going to modify your model to capture the effect of selection?
Actually, no right back atya. That’s starting to look like a quote mine.
What I wrote was:
So you are still wrong, and YOUR preferred definition of sexual reproduction quite specifically includes yeast mating types (sexes).
DOES TOO!! However strongly a population is growing, if you start with two individuals you only have at most 4 alleles at any given locus. Most loci will have fewer. Then sampling effects alone will cause many of those to be lost, even in the 1st couple of generations, while observed mutation rates will not provide enough subsequent variation to account for the later population’s statistics. Perhaps you could demonstrate this ‘mutation effect’ with a spreadsheet.
Ah, so the variation comes from Mouse Cain’s wife, and the inhabitants of the mouse city he founded?
This is hilarious stuff, keep it up!
The time between a mouse being born and giving birth (generation time) is short, usually around 10 weeks. This means that you could find out for yourself, for very little cost, if Madam and Meve could have founded their race. Got a basement?
VIdeo the whole thing, do some sort of ‘summary’ and then wipe the smile of the doubters faces!
Out of interest, when did the characters in the story of Adam and Eve live Nonlin? About how long ago?
It’s kinda relevant because otherwise, why ‘read the story’ at all?
Nonlin.org,
My apologies, too, for suggesting your graphs were hand-drawn rather than the product of random-number generation and calculation. Thanks for providing more info.
Nonlin.org,
Yeah, that’s why people fall silent … 😁😁😁
Right-y-o, but first, let me compliment you on actually having done some modeling. I always appreciate people getting their hands dirty.
Now, let’s upgrade your model to include heritability. I suggest you change your first column to represent genotypic values, and have them propagate through the remaining nine generations. You can achieve this by doing the following:
1) Select the B3 cell like you did in your screenshot
2) Place a dollar-sign ($) before the letter A
3) Press Enter
4) Copy the B3 cell
5) Select the range B3:Jn, where n is the number of rows in your simulation (I can’t see how many you created)
6) Paste
What changed in your graph? Explain!
So Darwin did NOT mention “homogeneous populations in a constant environment”?
And Jock did NOT presuppose evolution by mentioning “allopatry”, right?
Then replace it by something you don’t observe on human time-scales. You are missing the point.
[closes eyes]
I refuse to discuss diversifying selection until you have learned some basic genetics. OK?
Not sure what you mean: “bad example of your “noise””
“Drift” means many things to many people. What do you mean here?
Also, what “selection”? The point was: this ‘divergence’ is what we WOULD see without ‘regression’. But we DON’T see this. Because of ‘regression’.
Did you read 9. :
“‘Drift’ is often invoked as a mechanism of ‘divergence of character’. This is wrong because ‘drift’ explains nothing as it is either aimless noise or due to adaptation and environmental change. Yet, as shown, adaptation is in no way ‘divergence of character’. In addition, the stable coexistence of several distinct variants within a homogenous population shows “gradualism”, “survival of the fittest”, and “natural selection” to be false because the alleles responsible are themselves distinct (no “gradualism”), they all “survive”, and neither is “selected” for or against.”?
You can’t do that. Say the measure is height. By changing A to $A in B:J, you’re now saying your height is dependent on that of your 10th gen grandfather and not at all on your father. This would be stupid.
The first column now represents the effect of having a particular genotype (the genotypic value). Since the genotype is transmitted unchanged through the lineage, the phenotype is dependent on the mother (not father; your model has no sexual reproduction).
1. Those were your words
2. This is not classical sexual reproduction. It’s a hybrid
3. How did you come up with two identical individuals? Prove 100% or even 99.994% in non-lab conditions.
Mother or father doesn’t matter. It doesn’t represent reality.
You didn’t receive genetic material from your parents? That’s sad to hear!
Look, I managed to recreate your “plumpudding” model.
Nonlin.org,
Must I spoon-feed you this?
A single spore, “M”, germinates, in the wild.
It grows and produces a ‘daughter’ bud, “D1”.
“D” grows and produces a bud of its own, “DD”.
M produces a second daughter bud, “D2”.
Here’s the fun bit: once M has produced D2, both M and D2 have switched mating type. They do this by changing 700bp out of 12 million.
Sooo, M is 100% identical to D2, and D1 is 100% identical to DD. (They are a paltry 99.994% identical across mating types.)
But when M mates with DD and D1 mates with D2, they simultaneously create two 100% identical diploids. And this is what usually happens in the wild. If you had bothered to check any of the links I included, you would already know all this.
Constructing the phenotype by summing an inherited genotypic value and a new environmental deviation each generation results in a stable model. I simulated 1000 individuals (let’s call them “nonlins”), that produce a single offspring. Discrete generations, no sex. Genotypic values and environmental deviations were generated by the function in Nonlin’s model ( I believe it is a random draw from a normal distribution with mean 0 and sd = 1).
And if we compare two consecutive generations we observe … regression to mediocrity.
Here are generation 3 and generation 4. The dashed line is the reference line with slope 1. The solid blue line is the regression line. So where did the magic regression to mediocrity enter the model? I sure didn’t put it in.
Nice story, but not what happens. Even in the “evolution” narrative, there was a first human couple: let’s call them “mitochondrial Eve and sperm Adam”. You’re lost.
Anyway. You started this nonsense by claiming (without a proof) a narrowing of the distribution and asking me to explain. Well if that’s your theory, you demonstrate and explain it.
Good point. Many invasive species including mice were occasionally introduced in isolated environments and, those that thrived are doing just as well as their mainland peers.
No worries.
wut